Своеобразие сократительной функции гладких М. позвоночных животных определяется не только особенностями их иннервации и гистологического строения, но и спецификой их химического состава: более низким содержанием контрактильных белков (актомиозина), макроэргических соединений, в частности АТФ, низкой АТФ-азной активностью миозина, наличием в них водорастворимой модификации актомиозина — тоноактомиозина и т. д.
Существенное значение для организма имеет способность гладких мышц изменять длину без повышения напряжения (наполнение полых органов, например мочевого пузыря, желудка и др.).
И. И. Иванов.
Скелетные мышцы человека, различные по форме, величине, положению, составляют свыше 40% массы его тела. При сокращении происходит укорочение М., которое может достигать 60% их длины; чем длиннее М. (самая длинная М. тела портняжная достигает 50 см), тем больше размах движении. Сокращение куполообразной М. (например, диафрагмы) обусловливает ее уплощение, сокращение кольцеобразных М. (сфинктеров) сопровождается сужением или закрытием отверстия. М. радиального направления, наоборот, вызывают при сокращении расширение отверстий. Если М. расположены между костными выступами и кожей, их сокращение обусловливает изменение кожного рельефа.
Все скелетные, или соматические (от греч. so'ma — тело), М. по топографо-анатомическому принципу могут быть разделены на М. головы, среди которых различают мимические и жевательные М., воздействующие на нижнюю челюсть, М. шеи, туловища и конечностей. М. туловища покрывают грудную клетку, составляют стенки брюшной полости, вследствие чего их делят на М. груди, живота и спины. Расчленённость скелета конечностей служит основанием для выделения соответствующих групп М.: для верхней конечности — это М. плечевого пояса, плеча, предплечья и кисти; для нижней конечности — М. тазового пояса, бедра, голени, стопы.
У человека около 500 М., связанных со скелетом. Среди них одни крупные (например, четырёхглавая М. бедра), другие — мелкие (например, короткие мышцы спины). Совместная работа М. выполняется по принципу синергизма, хотя отдельные функциональные группы М. при выполнении определенных движений работают как антагонисты. Так, спереди на плече находятся двуглавая и плечевая М., выполняющие сгибание предплечья в локтевом суставе, а сзади располагается трёхглавая М. плеча, сокращение которой вызывает противоположное движение — разгибание предплечья.
В суставах шаровидной формы происходят простые и сложные движения. Например, в тазобедренном суставе сгибание бедра вызывает пояснично-подвздошная М., разгибание — большая ягодичная. Бедро отводится при сокращении средней и малой ягодичных М., а приводится с помощью пяти М. медиальной группы бедра. По окружности тазобедренного сустава локализуются также М., которые обусловливают вращение бедра внутрь и кнаружи.
Наиболее мощные М. размещаются на туловище. Это М. спины — выпрямитель туловища, М. живота, составляющие у человека особую формацию — брюшной пресс. В связи с вертикальным положением тела М. нижней конечности человека стали более сильными, поскольку, кроме участия в локомоции, они обеспечивают опору тела. М. верхней конечности в процессе эволюции, напротив, сделались более ловкими, гарантирующими выполнение быстрых и точных движений.
На основе анализа пространственного положения и функциональной деятельности М. современная наука пользуется также следующим их объединением: группа М., осуществляющая движения туловища, головы и шеи; группа М., осуществляющая движения плечевого пояса и свободной верхней конечности; М. нижней конечности. В пределах этих групп выделяются более мелкие ансамбли.
В. В. Куприянов.
Патология мышц. Нарушения сократительной функции М. и их способности к развитию и поддержанию тонуса наблюдаются при гипертонии, инфаркте миокарда, миодистрофии, атонии матки, кишечника, мочевого пузыря, при различных формах параличей (например, после перенесённого полиомиелита) и др. Патологические изменения функций мышечных органов могут возникать в связи с нарушениями нервной или гуморальной регуляции, повреждениями отдельных М. или их участков (например, при инфаркте миокарда) и, наконец, на клеточном и субклеточном уровнях. При этом может иметь место нарушение обмена веществ (прежде всего ферментной системы регенерации макроэргических соединений — главным образом АТФ) или изменение белкового сократительного субстрата. Указанные изменения могут быть обусловлены недостаточным образованием мышечных белков на почве нарушения синтеза соответствующих информационных, или матричных, РНК, т. е. врождённых дефектов в структуре ДНК хромосомного аппарата клеток. Последняя группа заболеваний, т. о., относится к числу наследственных заболеваний.
Саркоплазматические белки скелетных и гладких М. представляют интерес не только с точки зрения возможного участия их в развитии вязкого последействия. Многие из них обладают ферментативной активностью и участвуют в клеточном метаболизме. При повреждении мышечных органов, например при инфаркте миокарда или нарушении проницаемости поверхностных мембран мышечных волокон, ферменты (креатинкиназа, лактатдегидрогеназа, альдолаза, аминотрансферазы и др.) могут выходить в кровь. Т. о., определение активности этих ферментов в плазме крови при ряде заболеваний (инфаркт миокарда, миопатии и др.) представляет серьёзный клинический интерес.
Лит.: Энгельгардт В. А., Ферментативные и механические свойства белков мышц, «Успехи современной биологии», 1941, т. 14, в. 2; Сент-Джиордьи А., О мышечной деятельности, пер. с англ., М., 1947; Иванов И. И., Юрьев В. А., Биохимия и патобиохимия мышц, Л., 1961; Поглазов Б. Ф., Структура и функции сократительных белков, М., 1965; Хайаши Т., Как клетки движутся, в кн.: Живая клетка, пер. с англ., 2 изд., М., 1966; Хаксли Г., Механизм мышечного сокращения, в сб.: Молекулы и клетки, пер. с англ., в. 2, М., 1967; Смит Д., Летательные мышцы насекомых, там же; Бендолл Дж., Мышцы, молекулы и движение, пер. с англ., М., 1970; Арронет Н. И., Мышечные и клеточные сократительные (двигательные) модели, Л., 1971; Лёви А., Сикевиц Ф., Структура и функции клетки, пер. с англ., М., 1971; Иванов И. И., Некоторые актуальные проблемы эволюционной биохимии мышц, «Журнал эволюционной биохимии и физиологии» 1972, т. 8, № 3; Gibbons I. R., The biochemistry of motility, «Annual Review of Biochemistry», 1968, v. 37, р. 521.
И. И. Иванов.

Рис. 4. Электронная микрофотография гладкомышечного волокна. Видны актиновые нити (показаны стрелками).
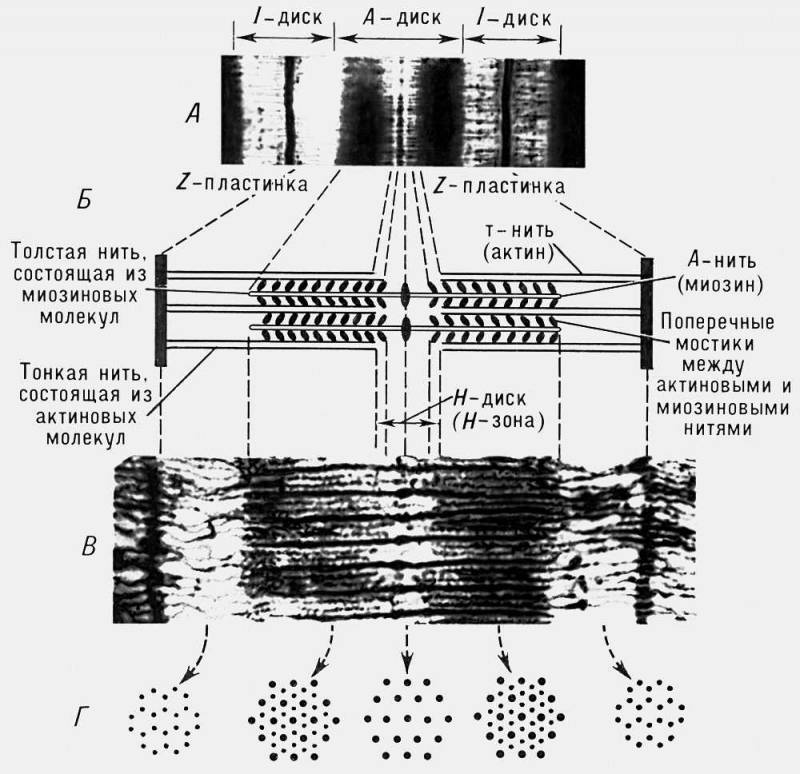
Рис. 3. Строение саркомера поперечнополосатого мышечного волокна: А — электронная микрофотография (малое увеличение), на которой четко видна структура саркомера; Б — схема саркомера; В — электронная микрофотография с высокой разрешающей способностью; Г — поперечное сечение саркомера на различных уровнях, видно положение толстых и тонких нитей в различных участках покоящегося саркомера (по Х. Хаксли).
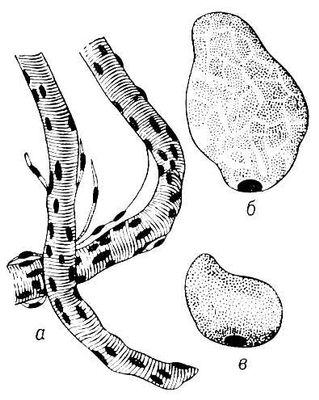
Рис. 1. Поперечнополосатые мышечные волокна человека: а — отрезки двух волокон; б — поперечный разрез волокна, миофибриллы образуют скопления, разграниченные саркоплазмой; в — поперечный разрез волокна с равномерным распределением миофибрилл.
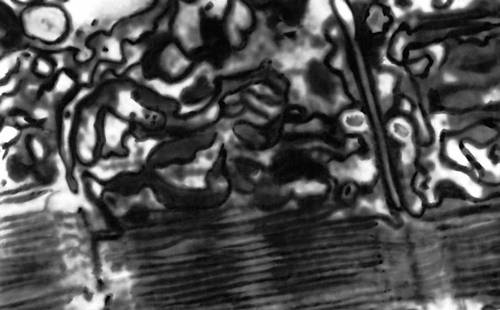
Рис. 2. Электронная микрофотография саркоплазматической сети и Т-системы мышечного волокна.