Среди металлов, входящих в состав организмов и преимущественно растений, калий наиболее распространенный и обильный элемент. А. И. Перельман высказал предположение, что биофильное поведение калия связано с его радиоактивностью в историческом аспекте. На заре своего развития жизнь, полностью еще не освоив механизм фотосинтеза, нуждалась в источниках энергии. Усваивая калий, первичные организмы получали не только вещество с нужными химическими свойствами, но и дополнительный источник свободной энергии. Это могло определить более интенсивное биологическое усвоение, которое закреплялось наследственностью. Поглощение калия передавалось и более высокоорганизованным формам жизни, для которых радиоактивность уже не имела существенного значения. Однако установившаяся физиологическая роль калия заставляла поглощать его в повышенных количествах.
Основным событием при зарождении первых организмов было образование спиральных молекул ДНК, что в УСЛОВИЯХ обилия органических веществ могло быть относительно быстрым процессом. Однако, по-видимому, возник не один организм, а живое вещество. И лишь значительно позже оно разделилось на индивидуальные сферические формы, ставшие родоначальниками организмов.
В дальнейшем в живом веществе происходили процессы усложнения. Произошло качественное изменение в эволюции живой материи, связанное с точностью воспроизведения нуклеиновых кислот как кодирующего процесса синтеза белков, которые значительно превосходили остальные органические соединения по своим биокаталитическим свойствам.
В процессе размножения новые организмы занимали все пространство, пригодное для жизни, что явилось важным условием завершения формирования биосферы в целом. В. И. Вернадский выдвинул принцип постоянства биомассы живого вещества, распространив его на всю историю планеты. Этот принцип был и остается глубоким научным обобщением. Однако следует подчеркнуть, что он имеет относительное значение. Величайшая напряженность жизни, выражающаяся в высоких темпах размножения мельчайших организмов, приводит к планетарному равновесию между естественной продукцией живого вещества и его разложением. Поэтому сейчас правильнее говорить о вековой тенденции к установлению постоянства биомассы для определенных, может быть даже значительных, интервалов геологического времени.
По данным молекулярной биологии, древнейшие микробы были представлены гетеротрофными организмами, которые размножались в среде с обильными органическими и минеральными питательными веществами. Эти питательные вещества включали по крайней мере рибозу, дезоксирибозу, фосфат, пурины и их предшественников, пиримидины, разнообразные «белковые» и «небелковые» аминокислоты. На ранних стадиях развития Земли фосфаты Na, К, Са имелись, вероятно, в достаточном количестве, как продукты выветривания первых горных пород. Кроме того, в качестве пищи могли быть использованы многие неизвестные или не идентифицированные соединения, в том числе некоторые смолообразные длинные полимеры.
Для первых организмов характерным был процесс ферментативного превращения органических веществ — брожение, где акцепторами электронов были другие органические вещества. Осуществление таких превращений в промежуточном обмене едва ли не во всех организмах служит аргументом в пользу древности этих процессов.
В ранней гетеротрофной биосфере Земли вскоре зародились организмы, способные поглощать углекислый газ, используя энергию солнечных лучей. По Л. Маргелис, биосинтетическая фиксация углекислого газа, столь обильного в первичной атмосфере Земли, происходила тремя способами.
Первая, наиболее примитивная фиксация была свойственна большой группе микроорганизмов, не чувствительных к видимому свету. Вторая возникла при участии фосфоенолпируват-карбоксилазы, которая наблюдается у анаэробных фотосинтезирующих бактерий. Третья фиксация СО2 совершалась при участии рибилозобиофосфат-карбоксилазы. Она присуща многим аэробным организмам и типична для большинства фотосинтетиков и хемоавтотрофов. Почти одновременно выработалась фиксация атмосферного азота. Это идущий с затратой энергии анаэробный процесс, обнаруженный только у прокариот.
Фотосинтетические пигментные системы образовались у прокариот еще до того, как последние в результате симбиоза стали пластидами эукариот. Можно полагать, что фотосинтез с выделением свободного кислорода возник первоначально вовсе не у зеленых растений, а у выделяющих его фотосинтезирующих бактерий и синезеленых водорослей.
Развитие биосферы Земли можно рассматривать как последовательную смену трех этапов (рис. 13). Первый этап — восстановительный — начался еще в космических условиях и завершился на Земле появлением гетеротрофной биосферы. Для первого этапа характерно появление малых сферических анаэробов (рис. 13, а). Присутствуют только следы свободного кислорода. Ранний способ фотосинтеза был, по существу, анаэробным. Развилась фиксация азота, поскольку часть ультрафиолетовой радиации проникала через атмосферу и быстро разлагала присутствующий аммиак.
Второй этап — слабоокислительный — отмечен появлением фотосинтеза. Он продолжался до завершения осадконакопления полосчатых железистых формаций докембрия. Аэробный фотосинтез начался предками цианобактерий. Кислород производился организмами, строящими строматолиты (рис. 13, б). Но кислород мало накапливался в атмосфере, так как реагировал с железом, растворенным в воде. При этом окислы железа осаждались, образуя полосчатые железистые формации докембрия. Только когда океан освободился от железа и других поливалентных металлов, концентрация кислорода начала возрастать по направлению к современному уровню.
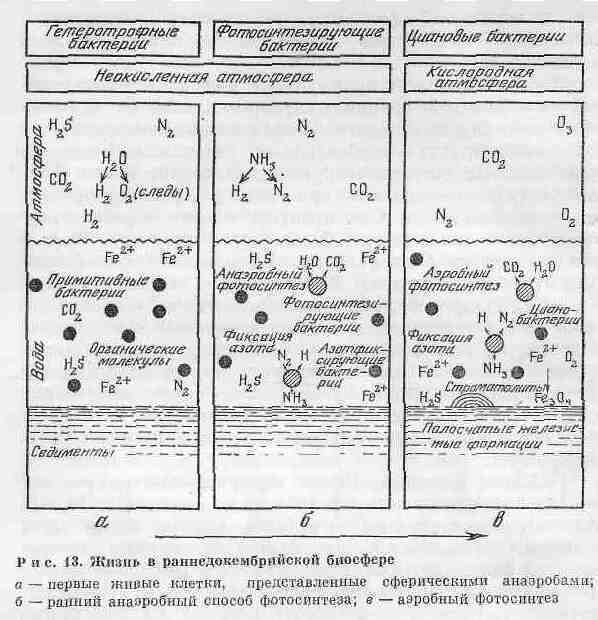
Третий этап характеризуется развитием окислительной фотоавтотрофной биосферы. Он начался с завершения отложений полосчатых железистых кварцитов около 1800 млн лет назад, в эпоху Карельско-Свекофенского орогенеза. Для этого этапа развития биосферы характерно наличие такого количества свободного кислорода, которого достаточно для появления и развития животных, потребляющих его при дыхании.
Последние два этапа в развитии биосферы фиксированы в каменной летописи геологической истории. Первый этап — наиболее далекий и загадочный, и расшифровка его истории связана с решением основных проблем органической космохимии.
Некоторые организмы раннего докембрия, относящиеся к синезеленым водорослям и пианобактериям, мало изменились в ходе геологической истории. Можно полагать, что простейшие организмы обладали наиболее устойчивой персистентностью (от латинского persiste — упорствую). По существу, в течение всей истории Земли не было причин для того, чтобы некоторые морские микроорганизмы, в частности синезеленые водоросли и бактерии, сильно изменились.
Растения как типичные представители фотоавтотрофных организмов нашей планеты возникли в ходе длительной эволюции, которая ведет свое начало от примитивных обитателей освещенной зоны моря — планктонных и бентосных прокариот. Сопоставляя палеонтологические данные с данными сравнительной морфологии и физиологии ныне живущих растений, можно в общем виде наметить следующую хронологическую последовательность их появления и развития:
1) бактерии и синезеленые водоросли (прокариоты);
2) водоросли циановые, зеленые, бурые, красные и др. (эукариоты, как и все последующие организмы);
3) мхи и печеночники;
4) папоротники, хвощи, плауны, семенные папоротники;
5) голосеменные (включая шишковые растения);
6) покрытосеменные, или цветковые, растения.
Бактерии и синезеленые водоросли обнаружены в наиболее древних сохранившихся отложениях докембрия, значительно позже появляются водоросли, и только в фанерозое мы встречаем пышное развитие высших растений: плауновых, хвощей, голосеменных и покрытосеменных.
В течение всего криптозоя в первичных водоемах в эвфотической зоне древних морей развивались преимущественно одноклеточные организмы — водоросли различного типа.