Отверстие замка, видимо, имеет форму чаши строго определенного объема. Молекулы с увеличенной головкой работали хуже. Укрупнить головку несложно, заменив метиловые группы
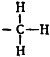
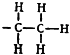

Ацетилхолин, как двуглавый орел, оказался с двумя головками. Расстояние между ними известно. В молекуле ацетилхолина атом азота отстоит от атома кислорода на 4,7 ангстрема. Если увеличить это расстояние, вставив в цепочку молекулы дополнительно один, два или больше атомов углерода, ключ не войдет в замочную скважину и двери не откроются.
Зная размер, отмычку подобрать нетрудно. Азот можно заменить любым другим атомом, лишь бы он нес положительный заряд и был отделен от атома кислорода двумя атомами углерода. Главное – размер. Атомы углерода тоже могут быть заменены. Головка этой стороны ключа имеет электрический заряд: третий от азота атом углерода несет положительный заряд, а один из атомов кислорода – отрицательный.
Ну хорошо, двери соседнего фасада открыты, что же дальше? Кто в них должен пройти? Оказывается, не сам медиатор (он всего лишь ключ), а положительно заряженный ион натрия.
Концентрация ионов в клетке и окружающей ее среде неодинакова. Внутри клетки много калия, а снаружи много натрия и хлора. Неравномерное распределение ионов приводит к тому, что внутри клетка заряжена отрицательно, а снаружи положительно. Когда медиатор открывает дверь, в нее устремляются малые катионы: натрий, калий и кальций. Более крупные не проходят, дверной проем для них слишком мал. Отрицательно заряженные ионы дверь не пропускает. Видимо, дверные косяки заряжены отрицательно, а одноименные заряды, как известно, отталкиваются.
Энергичнее всех движется натрий. Сзади его подталкивают силы диффузии, спереди притягивает отрицательный заряд. Навстречу натрию идет калий. Выбираться ему из клетки трудно. Правда, силы диффузии подталкивают, но сзади держит отрицательный заряд, а спереди отталкивает положительный. Поэтому калий движется значительно медленнее натрия. Перемещение ионов приводит к изменению наружного и внутреннего зарядов клетки, они начинают уравниваться.
Это можно сравнить с коротким замыканием. Если оно произошло, автоматически открываются новые двери, специально предназначенные для иона натрия. Он в еще больших количествах устремляется внутрь клетки. В результате внутренний заряд клетки становится положительным, а наружный из-за убыли положительно заряженного иона – отрицательным. Это приводит к возникновению потенциала действия, то есть к возбуждению клетки.
В тормозных синапсах дверь открывается только для определенного иона. Если открыта калиевая дверь, он устремляется из клетки наружу. Происходит не уравнивание зарядов, а усиление прежних, и клетка становится невозбудимой. В других случаях медиатор открывает двери для отрицательно заряженного иона хлора, сосредоточенного главным образом снаружи. Устремившись в открытую дверь, хлор также увеличивает отрицательный заряд внутри клетки и положительный – снаружи, делая клетку невозбудимой.
О самих дверях известно меньше, чем о запорах. Предполагают, что клеточная мембрана состоит из четырех слоев: два внутренних слоя образуются липидами, а два наружных – белками. Вероятно, липидный слой не сплошной. В местах разрыва липидной оболочки молекулы белка могут проникать – внутрь, двигаясь навстречу друг другу по его краям, пока не встретятся. В этом месте сначала должна появиться вмятина, а затем и дырочка. Электрические заряды ключа-медиатора являются толчком, вызывающим дырообразование.
Изучение роли медиаторов позволило выяснить, что же ломает кураре: замок или ключ. Оказалось, ни то и ни другое. Молекула курарина имеет сродство к холинорецептору, то есть к замку. Оно, как ключ, вставляется в замочную скважину, однако открыть дверь не может. У нее нет электрического заряда. Настоящий ключ (ацетилхолин) в заткнутую замочную скважину попасть не может. Двери остаются запертыми, возникает паралич.
Нарушить работу синапса можно многими способами. Удалось синтезировать вещества, которые препятствуют холину, образовавшемуся в результате распада ацетилхолина, всасываться обратно в нервное окончание. Тогда, если мозг беспрерывно шлет приказы и ацетилхолин постоянно выделяется в синаптическую щель, запасы его могут вскоре иссякнуть, а синтез прекратится из-за отсутствия исходного материала. Можно «подсунуть» в нервное окончание вещество, из которого вместо ацетилхолина синтезировались бы похожие молекулы, но не обладающие его активностью. В обоих случаях наступит паралич.
Кураре не получил применения в медицинской практике, но фармакологи отыскали немало соединений, которые можно использовать для устранения судорожных сокращений мышц. В их числе обнаружились вещества, отлично блокирующие холинорецептор, хотя их строение сильно отличалось от структуры ацетилхолина. Их молекулы были значительно крупнее, и вместо одной они имели две отрицательно заряженные головки. У наиболее эффективных веществ атомы азота были разделены 10 или 16 атомами углерода, то есть расстояние между ними равнялось соответственно 14 и 20 ангстремам. Загадка не из легких.
Наиболее достоверно предположение, что двери или, во всяком случае, замочные скважины не разбросаны как попало, а сгруппированы по четыре. Тогда молекула курареподобного вещества длиною 14 ангстрем будет иметь возможность, располагаясь вдоль любой из сторон квадрата, блокировать сразу два холинорецептора. В этом случае, чтобы подавить все холинорецепторы, нужно вдвое меньше молекул, а эффект окажется более надежным, так как ключ прочнее удерживается в двух замочных скважинах сразу. Понятен и эффект действия более длинных молекул. Они, располагаясь по диагоналям квадрата, также могут одновременно заткнуть две замочные скважины, блокировав два холинорецептора. Возможно, квартеты замочных скважин тоже сгруппированы и длинные молекулы курареподобных веществ могут блокировать две замочные скважины из разных квартетов. Вероятно, холинорецепторы функционально объединены, поэтому введение в одно из отверстий ключа повышает активность остальных трех замочных скважин, и они охотнее соединяются с ацетилхолином.
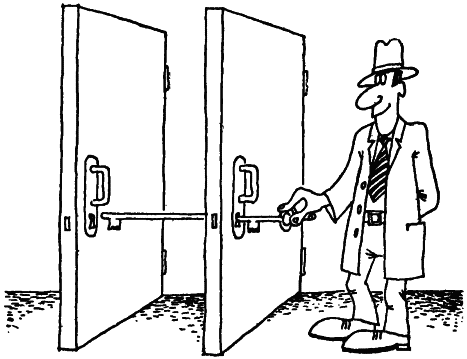
Расположение мышечных рецепторов было не всегда так строго упорядочено. У низших моллюсков и асцидий (одного из наиболее примитивных хордовых) замочные скважины разбросаны беспорядочно. Упорядочение холинорецепторов по два наблюдается у осьминогов, морских звезд и миног. Мышцы этих животных очень чувствительны к веществам, в которых атомы азота разделены 16 атомами углерода. У рыб обнаруживается активность и к препаратам с 10 атомами углерода. Значит, у них уже появляются квартеты холинорецепторов. А у птиц и млекопитающих отдельных пар, видимо, нет, и существуют только квартеты. Новорожденные малыши квартетов не имеют. У мышат, крысят, котят, щенят они формируются по мере роста.
Как медиаторы попадают в синаптическую щель, еще неясно. Существует предположение, что нервный импульс открывает очень немного дверей, но ключи, высыпавшиеся из них, используются в первую очередь для того, чтобы снаружи открыть остальные двери. Только теперь, когда ключей в синаптической щели оказывается достаточно много, они попадают в замочные скважины дверей на противоположном фасаде.