В отличие от других атмосферных газов кислород не может быть получен путем дегазации магмы, поэтому он отсутствовал в первичной атмосфере Земли, которая была восстановительной. Об этом свидетельствует наличие в древнейших, главным образом катархейских, породах легко окисляемых, но не окисленных (несмотря на пребывание на земной поверхности) веществ – таких, как графит (углерод), лазурит и т.д. Небольшие количества кислорода, образовывавшегося в результате фотодиссоциации молекул воды под действием жесткого ультрафиолетового излучения, полностью расходовались на окисление постоянно выделяемых вулканами аммиака, закиси углерода, метана и сероводорода. Однако с определенного момента в океанах начали образовываться так называемые полосчатые железные руды (джеспиллиты), представляющие собой чередование бедных и обогащенных окислами железа прослоев. Они формируются, когда двухвалентное (закисное) железо, выработавшееся в результате вулканической активности и растворенное в морской воде, реагирует в поверхностных слоях океана с молекулярным кислородом, окисляется до трехвалентного (окисного) и выпадает в виде нерастворимого осадка на дно. Таким образом, начало образования руд этого типа (состоящих в основном из гематита Fe2O3 и магнетита FeO · Fe2O3) означает появление на Земле достаточно мощного источника молекулярного кислорода, каковым могут быть только фотосинтезирующие организмы. Впервые джеспиллиты появляются все в той же формации Исуа; это может служить независимым подтверждением того, что содержащееся в ней углеродистое вещество действительно принадлежало фотосинтетикам (рис. 16).
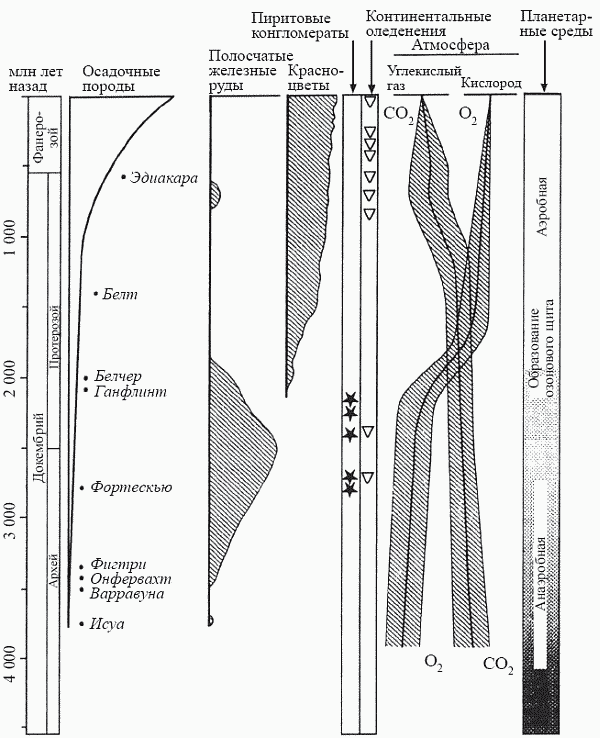
Источник-то кислорода возник, но мир еще на протяжении полутора миллиардов лет оставался анаэробным; об этом свидетельствует наличие в соответствующих отложениях конгломератов из пирита (FeS2). Зачастую они представляют собой гальку с отчетливыми следами обработки ее течением – это, кстати, первое свидетельство существования на Земле пресных вод. Но текучие воды не могут не быть хорошо аэрироваными, и то, что столь легко окисляемое вещество, как пирит, осталось неокисленным, свидетельствует о практически бескислородной атмосфере. Сообщества фотоавтотрофов формируют в это время своеобразные кислородные оазисы (термин П. Клауда) в бескислородной пустыне; их возможностей хватает на создание окислительных обстановок (и осаждение железа в окисной форме) лишь в своем непосредственном окружении.
Ситуация радикально изменилась около 2 млрд лет назад, когда процесс гравитационной дифференциации недр (см. главу 3) привел к тому, что бо?льшая часть железа перешла в ядро планеты и растворенное в морской воде закисное железо не могло уже возобновляться по мере выбывания из цикла. Фотоавтотрофы получили возможность завершить свою титаническую работу по окислению закисного железа и осаждению его в виде джеспиллитов; именно в это время возникли все крупнейшие месторождения железа, такие, как Курская магнитная аномалия. В дальнейшем руды этого типа уже не образовывались (за исключением очень краткого эпизода в самом конце докембрия – о нем речь пойдет в главе 6). Кислород, ранее полностью расходовавшийся на этот процесс, стал теперь мало-помалу насыщать атмосферу; с этого времени пиритовые конгломераты исчезли, а на смену донно-морским железным рудам пришли терригенные (имеющие наземное происхождение) красноцветы, т.е. процесс окисления железа начался и на суше (рис. 16). Яркий образ для описания этих событий придумал Г. А. Заварзин: он назвал их временем, когда «биосфера „вывернулась наизнанку“ за счет появления кислородной атмосферы как продукта обмена цианобактерий, и вместо кислородных «карманов» появились анаэробные «карманы» в местах разложения органики».
Именно в это время (1,9 млрд лет назад) в канадской формации Ганфлинт впервые появляются звездчатые образования, полностью идентичные тем, что образует ныне облигатно-аэробная марганцевоосаждающая бактерия Metallogenium. Без кислорода окисление железа и марганца не идет, и образуемые этой бактерией металлические кристаллы в виде характерных «паучков» возникают только в сильно окислительной обстановке. Это должно означать, что в тот момент содержание кислорода в атмосфере уже достигло величины как минимум в 1% от современного (точка Пастера). Именно с этой пороговой концентрации становится «экономически оправданным» налаживание процесса кислородного дыхания, в ходе которого из каждой молекулы глюкозы можно будет получать 38 энергетических единиц (молекул АТФ) вместо двух, образующихся при бескислородном брожении. С другой стороны, в атмосфере начинает возникать озоновый слой, преграждающий путь смертоносному ультрафиолету, что ведет к колоссальному расширению спектра пригодных для жизни местообитаний.
Одним из симптомов произошедшей «кислородной революции» стало то, что у автотрофов возникли серьезные проблемы с азот-фиксацией, т.е. переводом атмосферного азота в усвояемую, ионную форму. Азотфиксация возможна только в бескислородных условиях или при низком содержании кислорода, поэтому у автотрофов (таких, как цианобактерии), выделяющих кислород в процессе собственной жизнедеятельности, азотфиксация идет в специальных клетках с «герметической» утолщенной оболочкой – гетероцистах и акинетах. Акинеты впервые появляются в палеонтологической летописи как раз после «кислородной революции» – около 1,4 млрд лет назад.
Примерно к середине протерозоя (1,7–1,8 млрд лет назад) «кислородная революция» в целом завершается и мир становится аэробным (рис. 16). Впрочем, с точки зрения существ, составлявших тогдашнюю биосферу, этот процесс следовало бы назвать иначе: необратимое отравление кислородом атмосферы планеты. Смена анаэробных условий на аэробные не могла не вызвать катастрофических перемен в структуре тогдашних экосистем. В действительности «кислородная революция» есть не что иное, как первый в истории Земли глобальный экологический кризис.
Во всех предыдущих построениях мы имели дело лишь с геохимическими следствиями тех процессов, что происходили на Земле при предполагаемом нами участии живых существ. Но располагаем ли мы хоть какой-то информацией о самих этих существах? Можно ли их «пощупать пальцами»? Оказывается, можно. В последние десятилетия были разработаны специальные методы обработки осадочных горных пород, позволяющие выделять содержащиеся в них клеточные оболочки, а в некоторых случаях даже получать косвенную информацию о внутреннем строении этих клеток. К настоящему моменту в докембрийских породах обнаружено множество одноклеточных организмов, древнейшие из них – в местонахождениях Варравуна в Австралии (3,5 млрд лет) и Онфервахт в Южной Африке (3,4 млрд лет). Это оказались несколько видов цианобактерий (сине-зеленых водорослей), ничем особенно не отличающихся от современных; мы уже упоминали о том, что из пород возраста 3,1 млрд лет был выделен пигмент фикобилин, который используется при фотосинтезе современными цианобактериями (никем, кроме них).
Итак, в раннем докембрии существовал совершенно особый мир, формируемый прокариотными организмами – бактериями и циано-бактериями. Разделение живых существ на прокариоты и эукариоты (эти термины были введены в 1925 году Э. Шаттоном), основанное на наличии или отсутствии в их клетках оформленного ядра, теперь считают существенно более фундаментальным, чем, например, разделение на «животные» и «растения». Из многих их различий для нас сейчас важнее всего то, что среди прокариот неизвестны многоклеточные организмы. Есть нитчатые и пальмеллоидные[12] формы цианобактерий, однако уровень интеграции клеток в этих структурах – это все-таки уровень колонии, а не организма. А поскольку времени на всякого рода эксперименты у прокариот, как мы видели, было более чем достаточно (3,5 млрд лет), то приходится предположить, что по каким-то причинам на прокариотной основе многоклеточность не возникает в принципе. Может быть, дело в отсутствии у них центриолей и митотического веретена, без которого невозможно точно ориентировать в пространстве делящиеся клетки и формировать из них сколь-нибудь сложные ансамбли. Может быть – в свойствах их клеточной оболочки, препятствующих межклеточным взаимодействиям; это делает невозможным обмен веществами, так что клетки, находящиеся внутри организма и не имеющие прямого контакта с окружающей средой (что неизбежно при многоклеточности), не могли бы питаться и выводить продукты распада. Может быть – в чем-то еще. Как бы то ни было, такой путь повышения уровня организации, как многоклеточность, для прокариот оказался закрытым. Выход из этого тупика они нашли весьма оригинальный.
12
Пальмеллоидная структура выражается в образовании водорослями достаточно крупных (обычно прикрепленных к субстрату) слизистых тел, содержащих внутри многочисленные клетки; клетки объединяются в слизи чисто механически и плазматических связей не имеют.