Касаясь совершенства баланса между процессами синтеза и деструкции, осуществляемыми в мате, уместно вспомнить еще одно высказывание Вернадского, сделанное им в 1931 году: «Первое появление жизни при создании биосферы должно было произойти не в виде появления одного какого-либо организма, а в виде их совокупности, отвечающей геохимическим функциям жизни». Похоже, что и это предсказание теперь тоже находит подтверждение.
Каковы же были условия существования этих древнейших прокариотных экосистем? Они традиционно считались подводными (морскими) до тех пор, пока И. Н. Крылов и Г. А. Заварзин (1988) не выдвинули ряд аргументов в пользу наземного образования докембрийских строматолитов. Палеонтолог А. Г. Пономаренко (1993) задался таким вопросом: что такое вообще «море» и «суша» применительно к глубокому докембрию? Раньше это просто никому не приходило в голову, хотя постановка этого вопроса абсолютно правомерна. В отсутствие высшей наземной растительности скорость водной и ветровой эрозии должна превышать современную во много раз. Незащищенные наклонные субстраты при этом будут размываться очень быстро; ландшафт окажется состоящим из практически вертикальных скал и почти горизонтальных равнин, лишь незначительно превышающих уровень моря. Так что, по всей видимости, в те времена на окраинах континентов формировались обширные мелководные бассейны с постоянно меняющимся уровнем воды и без настоящей береговой линии. Именно в таком «вымершем» ландшафте (это не было ни сушей, ни морем в современном смысле) и процветали строматолитовые экосистемы.
Помимо бентосных прокариотных экосистем, представленных матами, все это время существовали и планктонные, состоящие из сферических одноклеточных водорослей – акритарх[14] – и шарообразных колоний, напоминающих современный Volvox. Первые планктонные организмы появляются в палеонтологической летописи 3,5 млрд лет назад – одновременно со строматолитами.
Именно в планктонных экосистемах в протерозое (около 1,9–2 млрд лет назад) произошло то, что стало ключевым для дальнейшей судьбы биосферы: в составе фитопланктона появились такие акритархи, которых считают первыми на Земле эукариотами. Заключение об их эукариотности первоначально основывалось на размере клеток (они превышают 60 мкм, тогда как практически все известные прокариоты со сферическими клетками меньше 10 мкм). Однако позже на Среднесибирском плоскогорье (местонахождение Мирое-диха) было собрано множество экземпляров уникальной сохранности, на которых удалось реконструировать процес ядерного деления. А недавно в одном из австралийских нефтяных месторождений сходного возраста (1,7 млрд лет) были обнаружены стеролы – вещества, которые синтезируются только клеточными ядрами.
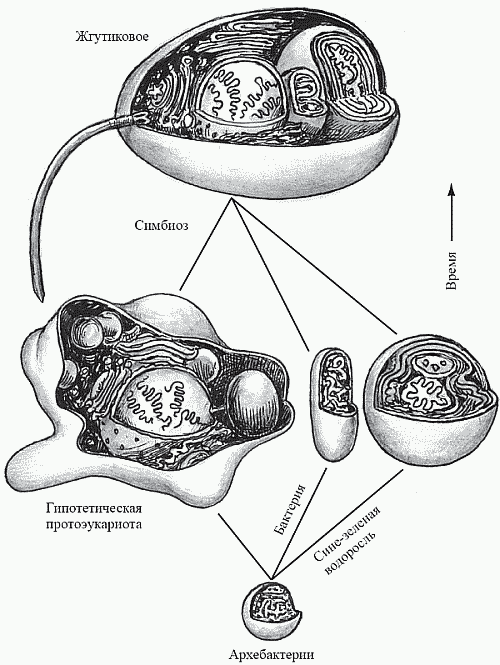
По поводу происхождения эукариот большинство исследователей придерживается гипотезы симбиогенеза (рис. 18). Идея о том, что растительная и животная клетки являют собой «симбиотический комплекс», была впервые высказана К. С. Мережковским (1909) и затем обоснована А. С. Фаминцыным (который ранее, в 1867 году, доказал симбиотическую природу лишайников); автором же концепции в ее современном виде считается Л. Маргулис. Концепция состоит в том, что органеллы, наличие которых отличает эукариотную клетку от прокариотной (митохондрии, хлоропласты и жгутики с базальным телом и микротрубочками), являются результатом эволюции некогда независимых прокариотных клеток, которые были захвачены клеткой-хозяином (тоже прокариотной) и не «съедены», а превращены ею в симбионтов. Предполагается, что роль клетки-хозяина выполняла крупная факультативно-анаэробная бактерия-гетеротроф. Проглотив однажды мелких аэробных бактерий-гетеротрофов, она начала использовать их в качестве «энергетических станций», перерабатывая с их помощью свою органику по более совершенной технологии (дыхание вместо брожения), что позволило ей получать из каждой молекулы глюкозы 38 молекул АТФ вместо двух. Союз был выгоден и для «рабов» – аэробов, получивших взамен гораздо более совершенный источник «топлива»: органику, добываемую крупным хищным хозяином. Далее к поверхности хозяина прикрепилась другая группа симбионтов – жгутикоподобные бактерии (Маргулис полагает, что это было нечто вроде современных спирохет), которые резко увеличили подвижность хозяина – в обмен на возможность «подсоединиться к единой энергосистеме». Поглотив затем подходящих мелких фотоавтотрофов – цианобактерий, хозяин обрел возможность получать органику не путем активного поиска ее во внешней среде, а просто, так сказать, посидев немного на солнышке. Этот переход до некоторой степени аналогичен происходившему в человеческой истории переходу от охоты и собирательства к скотоводству и земледелию.
Гипотеза выглядит достаточно фантастично, однако имеет серьезные обоснования. Аналогичные процессы происходят и в современном мире. Например, инфузория-туфелька может содержать в качестве «домашнего животного» зеленую водоросль хлореллу. Инфузория не трогает «домашнюю» хлореллу, но немедленно переваривает любую «дикую» клетку того же вида. Хлорелла же образует внутри хозяина строго фиксированное число клеток и через несколько поколений теряет способность к самостоятельному существованию.
С другой стороны, сходство митохондрий и хлоропластов с бактериями – не только внешнее (и те и другие – овальные тела, окруженные двойной мембраной, на складках внутреннего слоя которой локализованы пигменты с дыхательными или фотосинтетическими функциями). Эти органеллы имеют собственную ДНК, причем не линейную (как хромосомная ДНК самой эукариотной клетки), а кольцеобразную – такую же, как у настоящих бактерий. В момент клеточного деления митохондрии и хлоропласты не создаются клеткой заново, а размножаются сами, как настоящие одноклеточные организмы. При этом степень их самостоятельности такова, что они вполне способны существовать и размножаться и в чужих клетках: например, хлоропласты легко размножаются в курином яйце.
Сравнив с этих позиций эукариотную клетку с матом, можно заключить, что мы имеем дело со второй (и столь же успешной) попыткой прокариот создать высокоинтегрированную колонию, на этот раз не между различными клетками, а внутри одной из них. Похоже, что, не имея возможности выработать истиную многоклеточность, прокариоты выжали все, что можно, из симбиоза: раз нельзя наладить должную дифференцировку собственных клеток, то почему бы не использовать в качестве строительного материала клетки разных организмов? Не исключено, что «конструкторские разработки» по обоим «проектам» (мат и эукариотность) первые прокариоты начали одновременно. Такое предположение может хорошо объяснить тот парадоксальный результат, к которому пришли в последние годы К. Вёзе и ряд других исследователей: сопоставив нуклеотидные последовательности различных групп организмов методом молекулярных часов[15] , они заключили, что эукариоты должны были возникнуть столь же давно, как и прокариоты.
Скорее всего, различные варианты эукариотности, т.е. внутриклеточных колоний, возникали многократно (например, есть основания полагать, что красные водоросли, резко отличающиеся от всех прочих растений по множеству ключевых признаков, являются результатом такой «независимой эукариотизации» цианобактерий), однако все эти варианты, видимо, не имели в тогдашних условиях должного преимущества перед прокариотным типом организации и «не выходили в серию». Ведь эукариотам приходится платить за свою сложность весьма дорогую цену: они потеряли присущие прокариотам «бессмертие» (видовую неизменность на протяжении миллиардов лет) и «неуязвимость» (способность жить в кипятке или в ядерном реакторе, питаться любыми ядами или чистым водородом и т.д.); какой же смысл после этого выполнять в биосфере те функции, для которых достаточно и прокариот? Если компьютер нужен вам лишь для печатания текстов и игры в «Тетрис», то вы, надо думать, не станете покупать Пентиум со всеми «наворотами», а ограничитесь чем попроще.
14
Акритархи – это не какая-то таксономическая категория (как, например, эвгленовые или диатомовые водоросли), а собирательное название для микроорганизмов со стойкой, но неминерализованной оболочкой.
15
Метод датирования филогенетических событий, основанный на аксиоматичном утверждении, что нуклеиновым кислотам присуща практически постоянная (в геологическом масштабе времени) скорость эволюционно значимых замен молекулярных мономеров (нуклеотидов). Отсюда следует, что время, прошедшее с момента расхождения каждой пары ветвей родословного древа («сестринских групп») пропорционально числу молекулярных замен.