В рифее (1,4–1,2 млрд лет назад) эти нитчатые эукариотные формы достигли значительного разнообразия. Именно тогда в составе водорослевых лугов появились и первые макроскопические водоросли с пластинчатым, корковым и кожистым типами слоевища, а к венду (650 млн лет) основным компонентом этих сообществ становятся вендотении – лентовидные водоросли длиной до 15 см. Есть даже сообщение о находке каких-то пластинчатых водорослей в китайской формации Чанчен с возрастом 1,8 млрд лет; эта датировка нуждается в подтверждении, но не кажется нереальной. Дело в том, что таксономическая принадлежность всех этих форм недостаточно ясна, однако некоторые из них очень сходны с низшими красными водорослями – бангиевыми. А красные водоросли, как полагают многие исследователи, являются самой архаичной ветвью эукариот; возможно, они произошли в результате «независимой эукариотизации» цианобактерий.
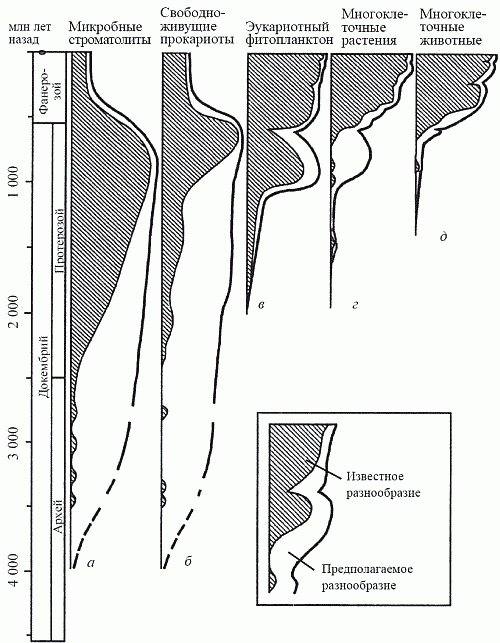
Животные и следы их жизнедеятельности (норки и следовые дорожки на поверхности осадка) достоверно появились в палеонтологической летописи лишь в конце протерозоя – около 800 млн лет назад. (Интересно, что водоросли с минерализованными слоевищами, для которых можно предполагать тот же уровень организации, что и у высших красных и бурых водорослей, появились еще позднее – в венде.) В чем же причина того, что истиная многоклеточность возникла так поздно? В 60-е годы прошлого века (вспомним главу 5) существовала гипотеза «кислородного контроля» Беркнера и Маршалла, согласно которой содержание кислорода в земной атмосфере вплоть до начала фанерозоя (540 млн лет назад) было ниже точки Пастера и не допускало существования более высоко организованных форм жизни, чем водоросли. Поскольку со временем установили, что точка Пастера в действительности была пройдена гораздо раньше – более чем за миллиард лет до времени появления первых многоклеточных, причинная связь между этими явлениями была отвергнута и о гипотезе «кислородного контроля» забыли. Как позже выяснилось, напрасно.
Однопроцентное содержание кислорода (имеется в виду 1% от его современного количества) – это тот критический минимум, ниже которого аэробный метаболизм принципиально невозможен; однако для жизнедеятельности макроскопических животных кислорода необходимо существенно больше. Б. Раннегар недавно провел специальные расчеты, из которых следует, что животным, составлявшим первую фауну многоклеточных – эдиакарскую (о ней речь впереди), кислорода требовалось не менее 6–10% от нынешнего уровня; это в том случае, если они имели развитую систему циркуляции, доставлявшую кислород к тканям. Если же такая система у них еще не развилась и они дышали за счет прямой диффузии (скорее всего, так и было), то требовалось для их жизнедеятельности гораздо больше кислорода (может быть, его содержание сопоставимо с нынешним). Итак, гипотеза «кислородного контроля» кажется вполне логичным объяснением появления макроскопических животных лишь в конце протерозоя – если принять более высокий критический порог, чем однопроцентный, предлагавшийся Беркнером и Маршаллом. Однако возможна ли проверка этой гипотезы? Мы помним (из главы 5), что можно по составу осадков отличить аэробные обстановки от анаэробных, но как определить количественные различия в содержании кислорода в былые эпохи?
Известно, что количество кислорода, создаваемого небиологическими процессами (фотолиз воды и т.д.), совершенно ничтожно; почти весь свободный кислород планеты создан фотосинтезирующими организмами. Однако живые существа не только производят кислород, но и потребляют его при дыхании. В биосфере осуществляется достаточно простая химическая реакция: nСО2+nH2O?(CH2O)n + nО2. «Читая» ее слева направо, мы получаем фотосинтез, а справа налево – дыхание (а также горение и гниение). Уровень содержания кислорода на планете стабилен потому, что прямая и обратная реакции взаимно уравновешиваются; так что если мы попытаемся увеличить содержание свободного кислорода в атмосфере путем простого наращивания объема фотосинтезирующего вещества, то из этой затеи ничего не выйдет. Сместить химическое равновесие, как вам должно быть известно из курса химии, можно, лишь выводя из сферы реакции один из ее продуктов. В нашем случае добиться увеличения выхода О2 можно, лишь необратимо изымая из нее восстановленный углерод в форме (CH2O)n или его производных.
Таким образом, производство кислорода биосферой начинает превалировать над потреблением этого газа (ею же), только если происходит захоронение в осадках неокисленного органического вещества. Этот вывод кажется достаточно парадоксальным и противоречащим расхожим представлениям. Так, например, природоохранная пропаганда вбила в голову широкой публики магическую фразу «Леса – это легкие планеты»; и мало кто дает себе труд задуматься над тем, что в действительности любое сбалансированное сообщество (в том числе тропический лес) потребляет ровно столько кислорода, сколько и производит. Если что и можно назвать «легкими планеты», так это болота, где как раз и идет процесс захоронения неокисленной органики. Поэтому, если мы установим, что в некий период геологической истории происходило интенсивное захоронение органического углерода, то мы вправе будем заключить, что в это время столь же интенсивно накапливался и кислород. А вот оценить темпы захоронения органического углерода в прошлые эпохи вполне возможно. Для этого существует специальный метод, основанный на изучении изотопного отношения 12C/13C в соответствующих осадках (органический углерод, участвовавший когда-либо в реакциях фотосинтеза, обогащен «легким» изотопом 12C).
Несколько лет назад на архипелаге Шпицберген была открыта уникальная по полноте последовательность позднепротерозойских осадков, отлагавшихся в период с 850 до 600 млн лет назад. Исследовав эти осадки на предмет изотопного отношения 12C/13C, Э. Нолль (1996) установил, что на протяжении всего этого времени темпы захоронения органического углерода оставались самыми высокими за всю историю Земли. Изучение позднепротерозойских осадков в других районах мира – в Канаде, Австралии и Южной Африке – подтвердило выводы Нолля. Итак, появлению макроскопической фауны предшествовало резкое увеличение количества свободного кислорода; вряд ли можно счесть это случайным совпадением (рис. 20).
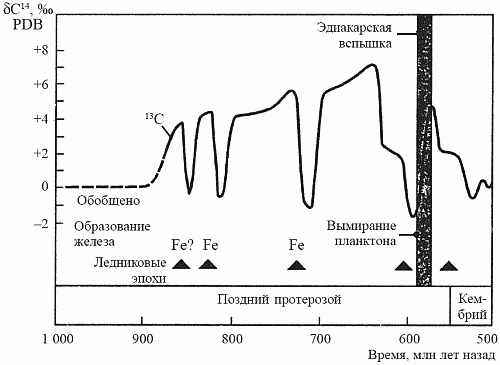
О том, что на Земле в это время действительно существенно возросло содержание кислорода, могут свидетельствовать и другие факты. Во-первых, в океанах вновь происходит осаждение джеспеллитов, прекратившееся около 1,8 млрд лет назад (см. рис. 16). Это может означать, что кислород впервые насытил глубоководные части океана, располагающиеся ниже фотической зоны, и при этом произошло осаждение еще сохранявшихся в тех глубинах запасов закисного железа. Во-вторых, это было время как минимум трех покровных оледенений, последнее из которых – Лапландское оледенение (600 млн лет назад) – было, по всей видимости, самым крупным за всю историю Земли. Одной из причин начала оледенений считают так называемый обратный парниковый эффект: когда в атмосфере планеты отношение О2/СО2 смещается в сторону кислорода, планета начинает гораздо хуже удерживать тепло, доставляемое центральным светилом.
Все это, вероятно, стало следствием событий, произошедших в биосфере около 1,2 млрд лет назад, когда начался постоянный рост численности и видового разнообразия эукариотного фитопланктона (закончился 600 млн лет назад). Известно, что в планктонных экосистемах детритная пищевая цепь развита очень слабо, а оборот вещества и энергии идет почти исключительно по пастбищной цепи; практически вся мертвая органика «проваливается» из экосистемы вниз, на дно водоема, где и утилизуется в тамошних детритных цепях. Однако донные детритные цепи протерозоя существуют в условиях сильного недостатка кислорода и потому обладают ограниченной «пропускной способностью». Начиная с некоторого момента они перестают справляться с тем нарастающим потоком органики, что поставляется сверху наращивающими свою продуктивность фитопланктонными сообществами. Другим источником органики могут быть лишенные в ту пору сколь-нибудь эффективных консументов («травоядных») водорослевые луга. В результате органический углерод начинает захораниваться в осадках на дне океана; оборотной стороной этого процесса является, как мы теперь знаем, накопление свободного кислорода, приводящее – через обратный парниковый эффект – к оледенению.