Давайте вернемся к графику, показывающему, как изменялось разнообразие фауны на протяжении фанерозоя (рис. 23). Обсуждаемые нами события знаменуют собой формирование новой биоты, которая сохраняет достаточное единство на протяжении всего палеозоя и большее разнообразие, чем прежняя, «кембрийская» – и в таксономическом, и в экологическом отношении. В современной экологии достаточно строго установлено, что смена эвтрофных условий на олиготрофные вызывает в экосистеме более «мелкую нарезку» экологических ниш и, следовательно, рост биологического разнообразия. Все это соответствует изложенным выше представлениям об «олиготрофизации» кембрийского океана за счет пеллетно-го транспорта.
В бентосе тем временем к обитателям раковин и жилых трубок добавляются существа с принципиально иными типами твердого скелета – внутренним (хордовые) и подвижным наружным (членистоногие). Интересно, что членистоногие (Arthropoda) появились позднее многих типов животного царства (во втором веке кембрия – атдабанском), однако сразу в составе практически всех своих подтипов и классов – и ракообразные, и хелицеровые, и вымершие трилобиты, и множество удивительных существ, точная систематическая принадлежность которых до сих пор неясна (рис. 24, в–д). Ситуация с кембрийскими членистоногими похожа на ту, что была с вендобионтами (см. главу 5): оказалось, что многие из них не могут быть отнесены к настоящим артроподам. Например, у аномалакариса (самого крупного животного того времени – 70 см) ротовые части состоят из четырех концентрически сходящихся лепестков, образующих нечто вроде зажимов цангового карандаша (рис. 24, г). Эти лепестки никоим образом не могут представлять собой видоизмененные конечности, что является одним из ключевых диагностических признаков типа членистоногих.
Вероятно, целый ряд неродственных между собой групп начинает в это время независимо приобретать отдельные признаки членистоногих, в результате чего они достигают артроподного уровня организации. Подробнее о процессах параллельной эволюции, подобных этой «артроподизации», мы поговорим позже (в главе 12). Пока же для нас существенно лишь само возникновение принципиально нового типа скелета, обладатели которого начинают играть ключевую роль в формирующихся экосистемах. Так, трилобиты становятся, по-видимому, основными потребителями того самого «упакованного» органического детрита, обогащающего донные осадки, и могут считаться настоящей эмблемой палеозоя.
К середине кембрия продуктивность и биологическое разнообразие достигают такого уровня, что у экосистем возникает нужда в «управляющем блоке» – верхних трофических уровнях, представленных специализированными хищниками-макрофагами. И вот начинается интереснейший процесс: борьба за этот «государственный военный заказ» между несколькими, самыми мощными, «корпорациями» – наиболее эволюционно продвинутыми группами животных. Из курса зоологии вам должно быть известно, что самыми высокоорганизованными среди первичноротых являются членистоногие и моллюски, а вершину эволюции вторичноротых представляют собой позвоночные. Именно эти группы и начинают соревнование за открывшуюся вакансию.
Первыми добились успеха членистоногие и близкие к ним арт-роподоподобные формы. Самым крупным хищником тех времен был аномалакарис; хищниками, как считают сейчас многие исследователи, были и самые первые трилобиты – лишь позднее эта группа перешла к детритофагии.[26] Однако вскоре вперед вырвались моллюски: появившиеся в конце кембрия головоногие (это были малоподвижные формы, родственные современному наутилусу, но с незакрученной конической раковиной, так называемые прямые головоногие) безраздельно господствовали в придонных слоях океана примерно до середины силура; на ордовик приходится пик видового разнообразия этой группы животных (около 150 родов). Хищники из числа членистоногих (ракообразные и водные хелицеровые – мечехвосты и ракоскорпионы) и позвоночных (бесчелюстные – родственники современных миног) оказываются «неконкурентоспособными» и занимают в экосистемах подчиненное положение.
Положение меняется в силуре, когда среди членистоногих появились крупноразмерные ракоскорпионы, а среди позвоночных – первые челюстноротые (настоящие рыбы). Судя по всему, обе эти группы возникли в опресненных прибрежных акваториях (типа лагун): здесь они были на первых порах избавлены от конкуренции с головоногими, которые избегают участков моря, «отравленных» стекающими с суши пресными водами. Именно в это время ракоскорпионы (например, Pterygotus) достигают длины свыше 2 м (а если с клешнями, то почти 3 м) и становятся властелинами силурийских морей – по крайней мере, их прибрежных участков (рис. 26). Прямые головоногие не выдерживают конкуренции, и их разнообразие заметно сокращается; попытки спастись за счет гигантизма (раковины Endoceras, например, достигали 4 м в длину, хотя, понятно, сам моллюск был много меньше) не имели успеха.
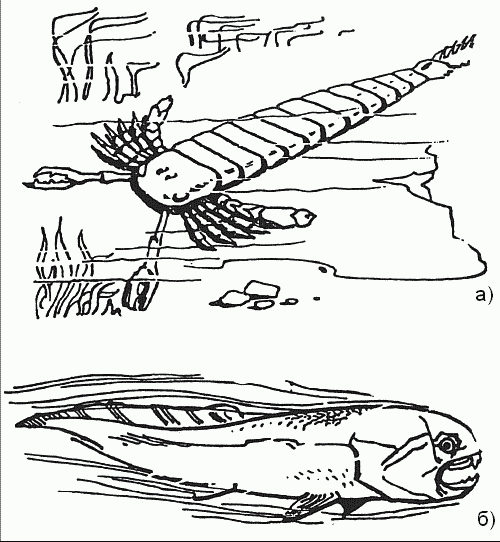
Однако торжество членистоногих (а именно ракоскорпионов) оказалось недолгим, и виною тому была третья «конкурирующая корпорация» – позвоночные. До сих пор те держались в тени и использовали лишь стратегию пассивной защиты – создание мощной наружной «брони»; таковы были панцирные бесчелюстные (остракодермы) и панцирные рыбы (плакодермы). В девоне одна из групп плакодерм – артродиры – отказалась от «бронирования корпуса» в обмен на скорость и перешла к активному хищничеству. Тогда-то и проявились все те (до поры скрытые) преимущества, что возникли ранее в результате превращения первой пары жаберных дуг в подвижные челюсти – великолепное средство нападения и защиты. Существа эти могли увеличивать размеры тела практически неограничено: такие артродиры, как динихтис, достигали 10 м в длину – одни из самых крупных рыб за всю историю (рис. 26), тогда как ракоскорпионы уже «уперлись» в физиологический предел размеров для членистоногих. В итоге артродиры стали для того времени фактически «абсолютным оружием», превосходя любого противника и по вооружению, и по скорости.
Членистоногие проиграли позвоночным эту схватку за крупный размерный класс вчистую: ничего подобного двухметровым силурийско-раннедевонским ракоскорпионам среди них не появится уже никогда. Отныне они будут методично осваивать малый и средний размерные классы (длина тела самых крупных членистоногих – камчатского краба и мечехвоста – не превышает 70 см), в чем и преуспеют: это самый процветающий ныне тип животного царства. А вот головоногие сумели тогда дать позвоночным достойный эволюционный ответ. Хотя численность придонных прямых головоногих продолжала сокращаться, суммарное разнообразие группы начиная с середины девона (эмский век) стремительно пошло вверх. К концу периода (фаменский век) оно достигло своего максимума (почти 200 родов) благодаря появлению свернутых головоногих, имеющих спирально закрученную раковину, – аммонитов.
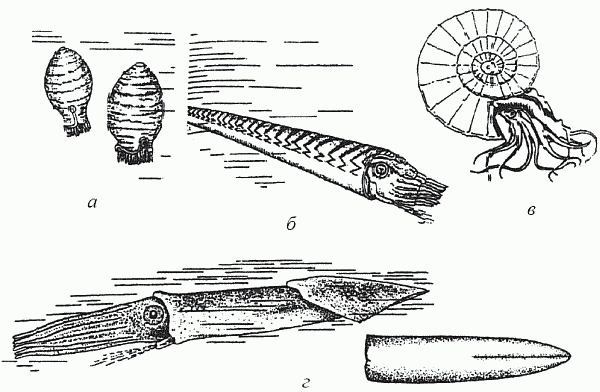
Прямые наутилоидеи обладали достаточно совершенным аппаратом регулирования плавучести (они меняли заполнение водой пустых камер раковины – так подводная лодка заполняет и опорожняет балластные цистерны), но вот с горизонтальными перемещениями у них были большие проблемы. Головоногие движутся реактивным способом, выталкивая воду из мантийной полости через воронку, направленную вперед и располагающуюся в районе головы. Первые наутилоидеи «висели» вниз головой в придонных слоях воды и могли лишь «падать» на свою добычу сверху (рис. 27, а). В дальнейшем они «завалили» раковину, сместив центр ее тяжести за счет неравномерного отложения карбоната кальция (на брюшной стороне больше, чем на спинной); вся конструкция в результате обрела способность к продольному движению (рис. 27, б). Однако эта конструкция принципиально неустойчива в движении, т. к. центр тяжести и точка приложения реактивной тяги (воронка) должны находиться на противоположных концах тела животного: малейшее отклонение от линии тяги – и движущийся моллюск начнет кувыркаться. А вот у свернутых головоногих центр тяжести их спиральной раковины почти совпадает с сифоном, что и обеспечивает устойчивое продольное движение. Именно поэтому свернутые головоногие стали первой группой хищников, освоивших толщу воды. (Здесь необходимо заметить, что рыбы, например, будут продолжать «ходить по дну» почти до середины мезозоя: лишь тогда костные рыбы «изобретут» плавательный пузырь, а среди хрящевых рыб появятся акулы современного типа, которые не «тонут» из-за того, что находятся в постоянном движении.) Видимо, именно освоение трехмерного пространства привело к возникновению у поздне-палеозойских головоногих самых совершенных (среди беспозвоночных) глаз и чрезвычайно усложненного мозга; то же произойдет позднее с птицами.
26
Описаны очень интересные нижнекембрийские следовые дорожки: по поверхности осадка идет общий след многощетинкового червя и трилобита, а потом след червя обрывается, и дальше ползет один трилобит (догнавший и съевший свою жертву).