Однако еще в середине прошлого века Т. Гексли, друг и сподвижник Ч. Дарвина, провел анатомическое сравнение четырех классов тетрапод, чтобы установить, кто чьим предком является. Глубокое сходство птиц и рептилий уже в те времена не вызывало особых сомнений; когда говорят, что «птицы – всего лишь продвинутая и сильно специализированная группа рептилий», это, в общем и целом, соответствует истине. Гексли обратил внимание и на менее очевидное, но весьма существенное сходство млекопитающих (маммалий) с амфибиями. Это сходство он видел в строении кожи (мягкой и богатой железами) и почек, выделяющих мочевину (у рептилий с птицами почки выделяют мочевую кислоту), а также в наличии двух затылочных мыщелков, которыми череп сочленяется с позвоночником (у рептилий с птицами – один). Но самым серьезным оказались для Гексли различия в строении кровеносной системы: у амфибий имеются две дуги аорты – правая и левая, у рептилий и птиц сохраняется только правая дуга, а у млекопитающих – только левая. Значит, «вывести» маммальную кровеносную систему из рептилийной (где левая дуга уже редуцирована) принципиально невозможно. На этом основании он заключил, что предками млекопитающих не могут быть рептилии (по крайней мере, нынеживущие) и маммалии должны вести свое происхождение прямо от амфибий.
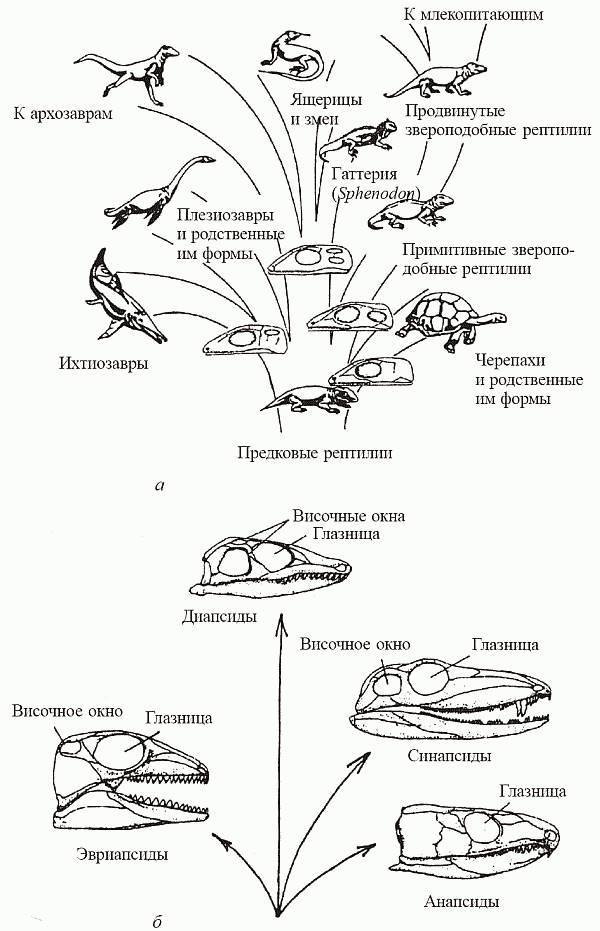
Так вот, по наиболее современным представлениям (они, как часто бывает в науке, могут считаться развитием на новом уровне взглядов Гексли), в нескольких (минимум в двух) эволюционных линиях амфибий независимо выработался синдром[55] рептилийных признаков и был достигнут рептилийный уровень организации. Выходит, «рептилии» – это категория не вертикальной, а горизонтальной классификации; это не таксон, члены которого связанны единством происхождения, а града – уровень организации, достигаемый тетраподами при утере ими облигатной связи с водой (рис. 39). Существование двух независимых эволюционных ветвей амниот – тероморфной (от греческого th?rion – зверь) и завроморфной (от греческого sauros – ящер), разошедшихся еще на уровне амфибий и венчаемых: первая – млекопитающими, а вторая – птицами и динозаврами, сейчас практически общепризнано[56] . Собственно говоря, все 300 млн лет существования наземных тетрапод – это история состязания теро-морфов с завроморфами, где эволюционный успех сопутствовал то одним, то другим.
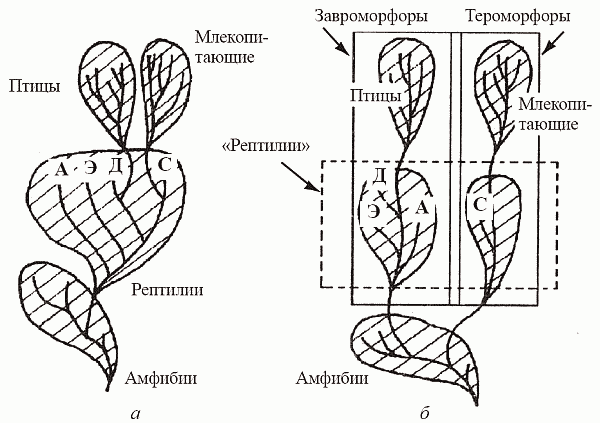
Первыми такого успеха добились тероморфы. Появившиеся еще в позднем карбоне пеликозавры – удивительные создания, многие из которых имели «парус» из остистых отростков позвонков (рис. 40, а) – составляли в ранней перми 70% всех амниот. Мощные клыки и непропорционально большая голова свидетельствуют о том, что пеликозавры были изначально ориентированы на питание крупной добычей; они были первыми хищными (в смысле плотоядными, а не насекомоядными) наземными существами. Тем интереснее то обстоятельство, что многие пеликозавры переходят к растительноядности – первый опыт фитофагии среди наземных позвоночных! Это чрезвычайно важно, ибо дефицит потребителей растительной массы в континентальных экосистемах должен был стать к тому времени просто угрожающим.
Основными фитофагами на суше являлись насекомые, но вначале они, как мы помним из предыдущей главы, только «снимали сливки» – потребляли одни высококалорийные генеративные части растений (пыльцу и семезачатки). В перми насекомые освоили питание соками растений, высасывая их при помощи хоботка из проводящих пучков (цикады и родственные им формы из отряда полужесткокрылых), а также древесиной, уже подвергшейся первичному грибному разложению (жуки). Однако питаться низкокалорийными живыми вегетативными тканями растений они стали лишь в мезозое. В карбоне (и даже еще раньше, в девоне) существовали другие потребители этого (прямо скажем, не особо привлекательного) ресурса: крупные двупарноногие многоножки; именно ими, как считают, оставлены погрызы на ископаемых листьях из палеозойских отложений. Пик разнообразия и обилия этих членистоногих пришелся на поздний карбон, а затем все они бесследно исчезли (возможно, разделив судьбу гигантских насекомых – см. предыдущую главу). Эту-то «вакансию» и заполнили первые позвоночные-фитофаги. Именно в раннепермских «пеликозавровых» сообществах впервые складывается нормальное для современности численное соотношение между растительноядными и хищниками (до того все тетраподы были либо рыбоядными, либо насекомоядными, т.е. хищниками, консументами второго порядка).
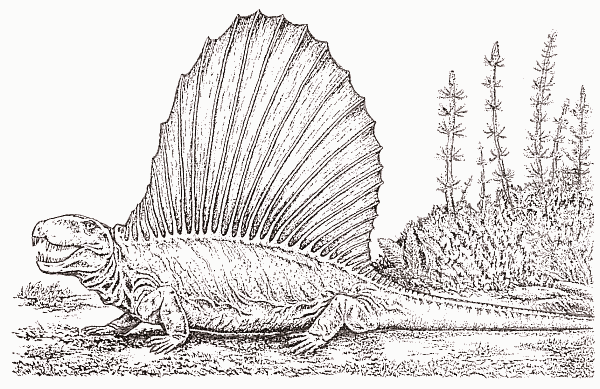
Мы уже говорили о том, что анатомия тероморфов просто-таки подталкивает их к различного рода эволюционным экспериментам с активной терморегуляцией. И хотя пеликозавры, без сомнения, были существами вполне «холоднокровными» (пойкилотермными) и метаболизм имели еще типично рептилийный, первые опыты по части температурного контроля начались уже на этом уровне. У многих из них возник «парус» – высокие остистые отростки позвоночника, заключенные в единую пластину из мягких тканей; в бороздах у основания отростков проходили крупные кровеносные сосуды. Парус, имевший огромную поверхность при малом объеме, служил, как сейчас считают, для быстрого поглощения или отдачи тепла телом. Расчеты показали, что особь пеликозавра массой 200 кг с парусом нагревается с 26 до 32°С за 80 мин (против 205 мин без паруса). По всей видимости, это позволяло пеликозаврам активизироваться по утрам значительно раньше добычи или других хищников сравнимого размера. С другой стороны, парус мог способствовать рассеиванию тепла в полуденное время, оберегая животное от перегрева. Характерно, что парус возникал как в хищных, так и в растительноядных группах пеликозавров, т.е. многократно и независимо. В дальнейшем сходные структуры изредка появлялись у динозавров (опять-таки как у хищных – спинозавр, так и у растительноядных – стегозавр), однако в целом такой способ терморегуляции оказался явно тупиковым.
По иному решили эту проблему более продвинутые тероморфы из отряда терапсид, появившиеся в поздней перми. Среди них были и специализированные растительноядные формы вроде дицинодонтов, достигавших размеров быка и имевших выступающие «моржовые» клыки в верхней челюсти (рис. 40, б), и крупные хищники – такие, как горгонопсиды, чем-то похожие на саблезубого тигра (рис. 40, в и г).
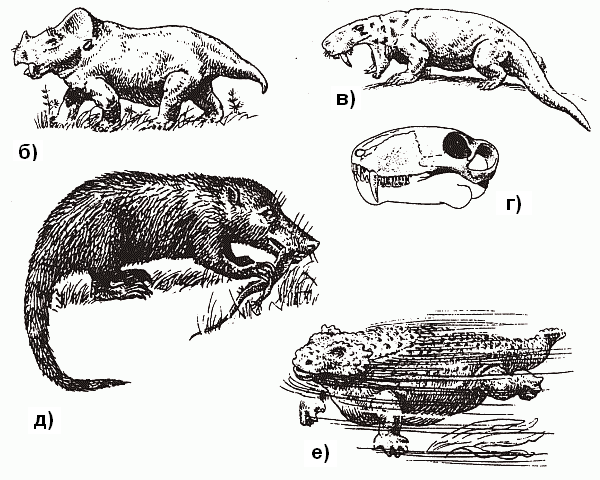
Еще недавно всех их считали просто «ящерами» (ну, зубы там дифференцированы, экая важность!) и именно в таком виде они обычно изображены в книжках по палеонтологии.
Однако сейчас полагают, что продвинутые терапсиды гораздо более походили на млекопитающих: доказано, что они, в отличие от прочих рептилий, имели развитое обоняние, слюнные железы и жесткие «усы»-вибри?ссы – осязательные волоски на морде (это можно установить исходя из того, как проходили поверхностные кровеносные сосуды черепа), а также «маммальное» строение поясов конечностей (об этом чуть ниже). Это были довольно «башковитые» создания – их коэффициент цефализации (отношение веса мозга к весу тела) заметно выше, чем у современных им завроморфов. Предполагают, что по уровню метаболизма эти животные не уступали наиболее примитивным млекопитающим – однопроходным (утконос и ехидна). Судя по всему, высшие терапсиды откладывали яйца, а детенышей выкармливали выделениями специальных желез – в точности, как однопроходные. Они были, скорее всего, покрыты шерстью (шерсть – это не что иное, как «расползшиеся» на всю поверхность тела и ставшие достаточно густыми осязательные вибрисы), а значит, являлись теплокровными: иначе зачем нужен теплоизолирующий покров?[57] Именно в одной из групп высших терапсид – цинодонтов (рис. 40, д) – в следующем, триасовом, периоде начался процесс, названный «параллельной маммализацией» (см. главу 11). Он, этот процесс, привел в конечном счете к возникновению настоящих млекопитающих.
55
Синдром – сочетание признаков (симптомов), имеющих общий механизм возникновения.
56
Палеонтолог М. Ф. Ивахненко (1980) привел серьезные доводы в пользу того, что и третья, «анапсидная», эволюционная линия тоже имеет независимые амфибийные корни. На этом основании он предложил выделить черепах, котилозавров и связанные с ними формы в отдельный класс – парарептилии. Кроме того, существует ряд доводов в пользу того, что амфибии, стоящие в основании соответствующих рептилийных стволов, ведут происхождение от разных групп кистеперых. Если эта гипотеза подтвердится, то окажется, что тероморфы и завроморфы разошлись даже не на «амфибийном», а еще на «рыбном» уровне!
57
О теплокровности высших терапсид может свидетельствовать и ряд косвенных признаков. В это время впервые появляются и подземные (роющие) животные, и ночные (с очень крупными глазами и бинокулярным зрением, как у лемура или совы), а такой образ жизни для пойкилотермного существа, нагревающегося «от солнышка», вряд ли возможен.