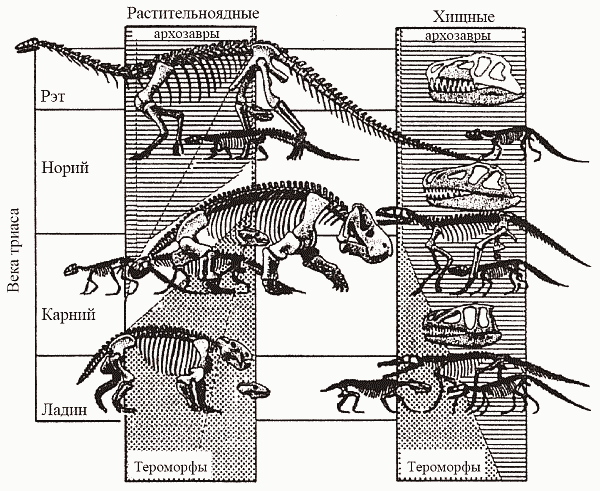
А что же другая, завроморфная, ветвь амниот? Они появились в том же позднем карбоне, что и тероморфы, но на протяжении всего палеозоя пребывали на вторых ролях. Правда, растительноядные анапсиды парейазавры (рис. 40, е) стали заметным элементом позднепермских экосистем, но в целом группа занимала явно подчиненное положение... И вот в начале мезозоя завроморфы берут реванш – да какой! В триасе представители тероморфной линии оказываются вытесненными на глубокую перифирию эволюционной сцены, а их место занимают завроморфы-диапсиды (рис. 41); последние к тому же осваивают такие ниши, которые до сих пор амниотам даже не снились – море и воздушное пространство. Триасовый упадок тероморфов, совсем уж было подошедших к тому, чтобы превратиться в маммалий, и внезапный расцвет «настоящих ящеров» – одна из самых интригующих загадок палеонтологии. В.А. Красилов удачно назвал эту ситуацию «мезозойским зигзагом». Упрощенные объяснения типа «иссушения климата» смотрятся тут абсолютно неубедительными – хотя бы потому, что пермь была временем даже более аридным, чем триас...
В триасе возникает целый ряд морских групп, эвриапсид: дельфиноподобные ихтиозавры, более похожие на ластоногих плезиозавров (всем хорошо известны эласмозавры с очень длинной шеей и маленькой головой – «змея, продетая сквозь черепаху», но в юре существовали и короткошеие плиозавры с черепом длиной до 3 м), а также менее известные нотозавры и плакодонты, несколько похожие на тюленей (рис. 42, а–г). Их появление в это время вполне логично следует из рептилийной физиологии. Говорят, что змея по вкусу больше всего похожа на цыпленка. Это чистая правда: у завроморфов (к коим принадлежат змеи) большая часть мускулатуры представляет собой «белое мясо», как грудные мышцы курицы. Белая мускулатура бедна (по сравнению с красной) миоглобином и митохондриями, и энергоснабжение мышечных сокращений в ней достигается в основном за счет гликолитического (анаэробного) расщепления глюкозы до молочной кислоты. Для этой реакции не нужен кислород, а скорость ее мало зависит от температуры, что особенно важно для животного, не обладающего гомойотермией. Однако (мы скажем в который раз!) за любое преимущество надо платить. Накапливающаяся в результате гликолиза молочная кислота – метаболический яд, и буквально через 1–2 мин интенсивной работы белая мускулатура практически теряет способность к дальнейшим сокращениям; самопроизвольный распад молочной кислоты (и восстановление работоспособности) требует нескольких часов. Белые мышцы принципиально не годятся для продолжительной работы.
Что касается богатых миоглобином красных мышц, составляющих основу мускулатуры маммалий, то тут ситуация иная. Они содержат огромное количество митохондрий (собственно, они и придают этой ткани красный цвет), и в них молочная кислота – лишь промежуточный продукт, который затем в митохондриях «дожигается» кислородом до углекислого газа и воды с выделением огромного количества энергии. Казалось бы, выгодно во всех отношениях (так что непонятно, зачем вообще кто-то сохраняет белые мышцы) – ан нет: реакции кислородного дыхания требуют повышенной температуры среды. Мало того, что для этого еще нужно выработать гомойотермию, так потом еще почти весь прибыток энергии придется потратить на собственный обогрев! Не зря млекопитающее потребляет (в покое) примерно в 10 раз больше пищи, чем рептилия равного с ним веса – а пищу эту, между прочим, надо еще поймать... В общем, «мамы всякие нужны, мамы всякие важны»: белая мускулатура хорошо (и энергетически дешево) работает «на рывке»[58] , а красная – при длительных постоянных нагрузках; животное с белыми мышцами – хороший «спринтер», а с красными – «стайер».
Вернемся к появлению в триасе разнообразных морских рептилий (кстати, и амфибии-лабиринтодонты тогда же сделали попытку освоить море). Многие исследователи (например, Р. Кэрролл) полагают, что переход к водному образу жизни происходит у низших амниот особенно легко именно в силу низкой интенсивности их метаболизма, способности выдерживать недостаток кислорода (из-за использования анаэробных процессов для работы мышц) и низкой температуры тела (не выше, чем у окружающей воды). Иными словами, рептилии исходно предрасположены к обитанию в водной среде.
Судя по наблюдениям над современными морскими игуанами, передвижение и питание в воде не требует от них структурной или физиологической перестройки; более того, при водной локомоции метаболические затраты этих ящериц составляют лишь четверть от тех, что потребны для перемещения по суше (благо тело в воде ничего не весит – по закону Архимеда).[59] Рептилии без проблем становятся вторичноводными всякий раз, когда жить в воде становится выгоднее с точки зрения обилия пищи и/или отсутствия врагов и конкурентов. Так, в юре возникли морские крокодилы (ноги их превратились в настоящие ласты, и имелся специальный хвостовой плавник, как у ихтиозавра), в раннем мелу – морские черепахи, близкие к нынеживущим, а в позднем мелу – мозазавры, исполинские (до 12 м длиной) морские ящерицы-вараны (рис. 42, д–е).
На суше ситуация складывается принципиально иначе, чем в море: тут белая мускулатура создает для завроморфов серьезные проблемы. Мы уже упоминали о различиях в строении поясов конечностей у тероморфов и завроморфов. Конечности могут быть расположены сбоку от тела (такое их положение называется латеральным) или подведены под него (парасагитально); в первом случае бедренная (или плечевая) кость направлена параллельно земле, а во втором – вертикально (рис. 43, а). Отсюда следуют различия в типе локомоции: рептилии «враскорячку» ползают на брюхе (отсюда происходит русское название группы – «пресмыкающиеся»), тогда как у маммалий тело высоко приподнято над землей, а конечность, двигаясь в плоскости, параллельной оси тела, способна делать широкий шаг (рис. 43, б). Весьма существенно то, что при парасагитальном положении конечностей (маммальный вариант) вес тела принимают на себя (продольно!) кости скелета конечностей, так что мышцам надо лишь не давать этим «подпоркам» отклоняться от вертикального положения; при латеральных же (рептилийных) конечностях тело «подвешено» на самих мышцах, и животное как бы постоянно выполняет упражнение «отжимание от пола» – удовольствие ниже среднего.

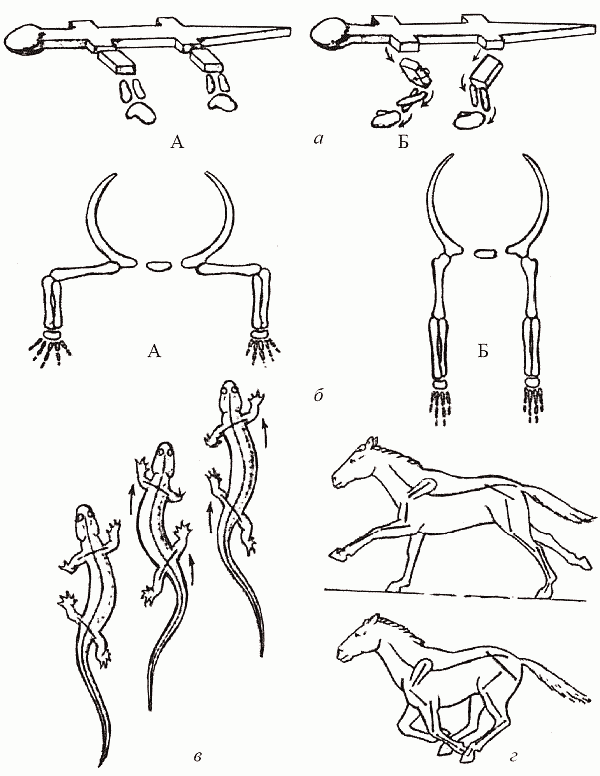
Рис. 43. Локомоция рептилий и маммалий: а – латеральное (А) и парасагитальное (Б) положение конечностей; б – постановка тела у рептилий (А) и маммалий (Б); в – «рептильный» шаг; г – «маммальный» шаг (по Шилову, 1994)
Казалось бы, маммальное строение поясов конечностей безусловно «лучше». В действительности же свои плюсы есть и в рептилийном варианте. Ведь если мускулатура по преимуществу белая, ориентированная «на рывок», то проще бо?льшую часть времени проводить, расслаблено лежа на брюхе, и лишь изредка совершать броски за добычей. При маммальной же постановке конечностей приходится постоянно поддерживать тело на весу; это, во-первых, практически исключает саму возможность использовать белую мускулатуру (которая «устает»), а во-вторых, заставляет тратить много энергии вхолостую (мы помним, что млекопитающее в покое расходует примерно в 10 раз больше энергии, чем рептилия равного размера). Однако когда все плюсы и минусы просуммированы, оказывается, что на суше для завроморфов (с их белой мускулатурой и латеральной конечностью) открыты лишь «профессии» подстерегающего хищника или пассивно защищенного – панцирем или слоем воды – крупного фитофага; остальные экологические ниши оказываются в распоряжении «теплых», активных, высокоподвижных тероморфов.