Появление и распространение покрытосеменных – одно из ключевых событий в истории биосферы, а вызваные им изменения сопоставимы по своим масштабам с появлением наземных (сосудистых) растений или животных с минеральным скелетом. Нынешний мир – ангиоспермный; голосеменным и споровым пришел в мелу такой же конец, как в позднем палеозое – водорослевым матам (при появлении сосудистой растительности). Ныне голосеменные растут лишь там, где им «позволяют»: сосна растет на песках и болоте, а лиственница – на вечной мерзлоте совсем не оттого, что им там хорошо, а оттого, что на эти неудобья «не позарился» никто из цветковых. В некоторых ландшафтах (в тропических лесах и в пустынях) голосеменных нет вовсе.[69]
Если задать ботанику вопрос «какие черты покрытосеменных привели их к эволюционному успеху?» – он наверняка начнет говорить о двойном оплодотворении, о триплоидном эндосперме и т.п.; все это, конечно, верно, но немножко не о том. «С точки зрения биосферы», наиболее существенны, пожалуй, трисвойства цветковых, позволившие им радикально изменить самую структуру мезозойских экосистем.
1. Склонность к неотении; в данном случае – к образованию травянистых форм, которые, по сути дела, есть древесные проростки, ставшие способными к размножению. Вспомним, что голосеменные «траву» не образуют вовсе (ни сейчас, ни в прошлом). Именно травянистые двудольные являются самыми агрессивными эксплерентами («сорными растениями»), мгновенно захватывающими места нарушений растительного покрова, а травянистые однодольные, склонные к образованию многолетней дернины, наиболее эффективно препятствуют эрозии.
2. Появление «мясистых» листьев с большим объемом фотосинтезирующей паренхимы, вызвавшее появление листогрызущих насекомых. До того насекомые потребляли лишь генеративные органы, содержимое сосудов, даже древесину, но только не листья: в листе голосеменного (даже такого, как гинкго) просто нечего есть. Ситуация меняется лишь с появлением цветковых – объеденные листья и мины (ходы, проедаемые личинками насекомых в толще листа) известны с самого начала позднего мела (сеноманский век). Обширная фауна листогрызущих насекомых сформировалась на покрытосеменных, а уже потом они перешли на голосеменные, папоротники и хвощи.
3. Энтомофилия цветков, открывающая покрытосеменным путь к сложной коэволюции с насекомыми. Сам цветок покрытосеменных, судя по всему, сформировался в процессе адаптации к насекомоопылению для защиты завязи от повреждений ее опылителями.
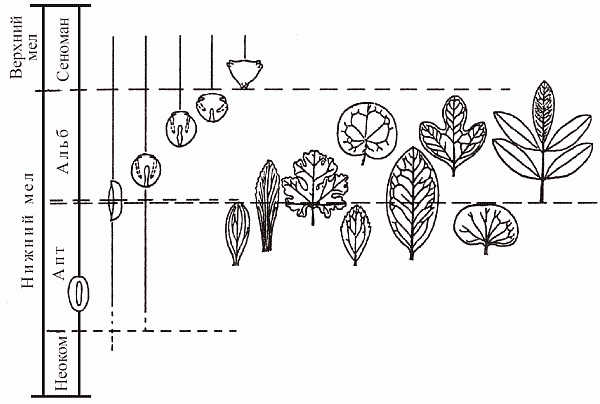
Ангиоспермы впервые появляются в палеонтологической летописи в начале раннего мела, в неокоме; на протяжении барремского и аптского веков они представлены единичными находками, в альбе идет стремительное нарастание их числа, а уже в самом начале позднего мела, в сеномане, цветковые составляют большую часть ископаемых флор, т.е. наступает кайнофит (рис. 50). Среднемеловая экспансия цветковых была столь стремительной (она фактически уложилась в ничтожный по геологическим меркам отрезок от 115 до 108 млн лет), что Дарвин в свое время в сердцах назвал этот эпизод палеонтологической истории «отвратительной тайной» (abominable mistery). Делались попытки объяснить это так: цветковые на самом-то деле появились на Земле давным-давно, минимум в юре; просто они обитали лишь в горах (не попадая по этой причине в палеонтологическую летопись), а в середине мела наконец заселили приводные местообитания, благоприятствующие попаданию в захоронения. Вообще такой «период скрытого существования» сбрасывать со счетов никак нельзя, но в данном случае эта гипотеза не проходит. Дело в том, что пыльца покрытосеменных (которая летает везде, и ее не «упрячешь» в горы) появляется в летописи одновременно с их макроостатками. По всем признакам, цветковые действительно возникли в начале мела, причем именно в приводных местообитаниях.
В 70-х годах советские палеоэнтомологи исследовали множество местонахождений меловых насекомых в Сибири и Монголии, «закрасив» тем самым наиболее крупное «белое пятно» в геологической истории класса. Оказалось, что у насекомых график, который отражает долю вымерших таксонов в фаунах разных времен, выглядит довольно неожиданно (рис. 51). Доля вымерших семейств мало меняется на протяжении большей части мезозоя; на границе юры и мела она составляет около 60%, затем, в течение раннего мела, стремительно падает от 60 до 20%, а в позднем мелу это падение резко замедляется (от 20 до 5%) и к началу кайнозоя кривая вновь «выходит на плато», только уже в районе 5%.
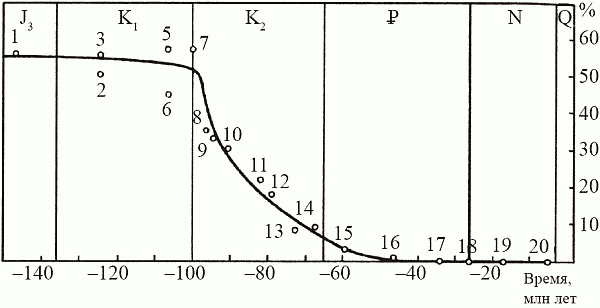
Иными словами, у насекомых самая резкая фаунистическая смена, связанная с массовым вымиранием мезозойских групп, приходится не на конец мела, а на его середину (в точности как у растений). На этом основании В. В. Жерихин (1978) предложил следующую модель среднемелового биоценотического кризиса.
Покрытосеменные возникли в начале мелового периода, первоначально как ценофобы – растения, не входящие в закономерные, сложившиеся в длительной коэволюции сукцессионные ряды.[70] В качестве «сорной» растительности они занимали свободные от других растений участки (речные отмели, береговые оползни, гари), которые соседствовали с гораздо более обширными участками, занятыми зрелыми сообществами мезофитной растительности. Покрытосеменные были исходно энтомофильными и смогли сформироваться лишь в окружении энтомофильных же голосеменных (таких, как беннеттиты, саговники, кейтониевые), используя уже сложившийся к тому времени комплекс их опылителей. Но если покрытосеменные оказались среди пионерных растений мезофита единственными энтомофилами (судя по всему, так оно и было), то это должно было дать им гигантские преимущества: именно для локально распределенных, не образующих сплошных зарослей пионеров насекомоопыление особенно выгодно; впрочем, возможно, имело место и прямое конкурентное вытеснение покрытосеменными энтомофильных голосеменных из пионерных сообществ – за счет более эффективного привлечения насекомых, лучшей защиты от повреждения завязи опылителями и т.д.
Основой стремительной среднемеловой экспансии цветковых стало то, что им удалось закрепиться в качестве нормальной пионерной растительности, для чего решающими факторами стали их исходная энтомофильность и наличие среди них травянистых форм (что во много раз убыстряет зарастание поврежденных участков). Покрытосеменные не пытались потеснить всю мезофитную растительность, что было абсолютно нереально; они «всего-навсего» конкурентно вытеснили прежних пионеров и тем самым блокировали все последующие стадии мезофитной экогенетической сукцессии. Разрушение существовавших в то время закономерных сукцессионных рядов (мезофитная растительность продолжала существовать там, где существовала, но она потеряла способность восстанавливаться после экзогенных нарушений) вызвало полный развал мезозойских наземных экосистем и массовое вымирание входивших в них животных; наиболее интенсивно эта деструкция шла в альбе (захватывая конец апта и начало сеномана).
В. В. Жерихин (1978) обратил внимание на любопытнейшую черту энтомофаун этого времени: многие семейства, характерные для мезозоя, обнаруживаются в них в последний раз, тогда как сколько-нибудь заметного появления новых групп не происходит. При этом в палеонтологической летописи вновь появляется целый ряд реликтовых для того времени (раннемезозойских и даже позднепалеозойских) групп. Это «всплывание реликтов» считают одним из наиболее ярких симптомов кризиса, охватившего среднемеловые сообщества: вымирание доминирующих раннемеловых групп (наиболее тесно связанных с существующей сукцессионной системой) создает экологический вакуум, который ненадолго заполняют древние формы, оттесненные к тому времени «на обочину жизни».
69
Единственные исключения – гнетум в первом случае, а также эфедра и вельвичия – во втором, но эти растения по своей жизненной форме являются имитаторами цветковых: у них есть широкий мягкий лист, ягоды и т.п. В некоторых новейших классификациях их даже стали включать в состав цветковых.
70
Под сукцессией здесь подразумевается не всякая смена растительности, а то, что С.М. Разумовский (1982) называет экогенетической сукцессией: жестко детерминированная последовательность растительных ассоциаций (сериальный ряд), каждая из которых видоизменяет состав почвы так, что он становится более благоприятен для ассоциации-последователя, нежели для нее самой, и приводит в конечном итоге к ассоциации-климаксу, в почвах которой достигнут «нулевой баланс» по органике. Климаксная ассоциация не имеет внутренних причин к смене и может существовать неопределенно долго – пока не будет разрушена извне (например, лесным пожаром). Пример экогенетической сукцессии для Подмосковья: зарастание гари иван-чаем (пионерная стадия), березняк, смена его ельником и далее дубравой (климаксом); цикл этот длится около 300 лет. Совокупность климакса с ведущими к нему сериальными рядами называют сукцессионной системой; ее можно рассматривать как гомеостат, который имеет равновесное состояние (климакс), а будучи же выведен из него внешними силами, возвращается в исходное положение при помощи ряда стандартизованных реакций (экогенетических смен)