Складывающиеся на совершенно новой основе раннекайнофитные сообщества первоначально были несбалансированными и неустойчивыми. Слабость биоценотической регуляции и обилие незанятых экологических ниш вели к очень быстрой, взрывной, эволюции различных групп организмов (из насекомых здесь можно отметить высших бабочек и мух, паразитических перепончатокрылых, а также общественных насекомых – муравьев и термитов), по мере же насыщения биоценозов темпы эволюции снижались. Сложные трофические цепи в новых кайнофитных сообществах сформировались очень быстро (листья, поврежденные минирующими бабочками, сами эти бабочки и их высокоспециализированные паразиты из числа наездников Braconidae появляются в летописи почти одновременно, в сеномане) и в дальнейшем менялись мало. «По экологическому облику и систематическому составу позднемеловые фауны значительно сильнее отличаются от раннемеловых, чем от кайнозойских, и могут рассматриваться как ранний, первоначально сильно обедненный, вариант этих последних, – заключает Жерихин (1980). – В сущности, по набору жизненных форм они имеют не так уж много отличий от современных».
Если вернуться к истокам среднемелового кризиса (самому возникновению покрытосеменных), то можно здесь обнаружить еще один интересный аспект, на который обратил внимание А. Г. Пономаренко (1997). Серьезные эволюционные изменения среди насекомых начинаются несколько раньше широкого распространения покрытосеменных – с конца юры, что, казалось бы, противоречит гипотезе о причинной связи между этими явлениями, но только на первый взгляд. Именно в это время начинается очередная «...зация» – ангиоспермизация голосеменных – и широко распространяются проангиоспермы (термин В. А. Красилова, 1989), т.е. голосеменные с отдельными чертами цветковых, например гнетовое Eoantha с крупными, явно энтомофильными «цветами» или беннеттит Manlaia, имитирующий осоку (рис. 52). «С точки зрения экосистемы», точное систематическое положение всех этих проангиоспермов не столь уж важно, т. к. все они могут в некоторой степени играть экологическую роль настоящих покрытосеменных.
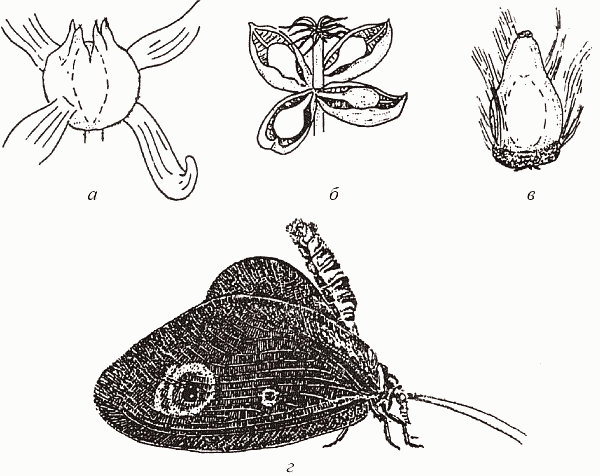
Неудивительно, что параллельно с энтомофильными голосеменными появляются антофильные насекомые; имитаторам цветковых в растительном мире сопутствуют столь же многочисленные имитаторы бабочек среди насекомых, например Kalligrammatidae из сетчатокрылых или Aneuretopsychidae из скорпионниц (рис. 52). Важнее же всего то, что здесь складывается система с положительной обратной связью: наличие у некоего беннеттита комплекса опыляющих его насекомых стимулирует появление энтомофилии у его соседей-голосеменных, что, в свою очередь, расширяет экологическую нишу этих насекомых-опылителей, и т.д. Каждый шаг на этом пути увеличивает вероятность следующего и делает появление покрытосеменных (т.е. растений с полным набором ангиоспермных признаков) совершенно неизбежным, а какая конкретно группа голосеменных даст начало «настоящим цветковым» (и кто из мекоптероидных насекомых породит «настоящих бабочек»), вообще-то говоря, не столь уж важно. Именно эти взаимно индуцируемые процессы Пономаренко и назвал «ангиоспермизацией мира».
Ангиоспермизация мира захватывает не только насекомых, но и все прочие компоненты континентальных экосистем. С начала мела на суше стремительно распространяются птицы (это особенно хорошо видно по укрупнению птерозавров – они полностью уходят в нишу крупных парителей) и териевые млекопитающие; в пресных водоемах настоящие костистые рыбы вытесняют ганоидов. После аптальбского кризиса мир обретает вполне знакомый нам облик; правда, в нем живут динозавры, да не просто живут, а вполне процветают на протяжении большей части позднего мела (судьба этой группы была рассмотрена нами в главе 11), и нет оснований напрямую связывать их вымирание с драматическими изменениями в морской биоте на границе мела и кайнозоя. Таким образом, мезозойско-кайнозойская смена на суше произошла много раньше, чем в море.
Ну а каковы же все-таки причины позднемелового Великого вымирания? Если не падение астероида и прочие «импакты», то что же? Давайте прежде всего попытаемся найти какие-нибудь общие черты для этого события и для другого морского вымирания – пермо-триасового. Оно было самым крупным в истории Земли: тогда в морских группах вымерло 50% семейств, 70% родов и более 90% видов (для сравнения: при мелкайнозойском вымирании исчезло менее четверти семейств). Помимо прямого исчезновения множества палеозойских групп (трилобиты, палеозойские кораллы – ругозы и табуляты), произошла радикальная смена структуры морских экосистем (роль основных рифостроителей перешла от мшанок к кораллам, роль донных фильтраторов – от брахиопод к двустворчатым моллюскам, и т.п.). Причины этой грандиозной фаунистической смены остаются загадкой. Стандартные объяснения вроде «глобального катастрофического вулканизма» или морской регрессии, резко сократившей площадь шельфов (основной зоны обитания морских организмов), кажутся просто придуманными «за неимением лучшего». Мне лично наиболее остроумной и логичной (хотя и не бесспорной) представляется гипотеза Х. Таппан, специалиста по фитопланктону.
В 1986 году Таппан опубликовала работу под названием Phytoplancton: below the salt at the global table («Фитопланктон: за солью на планетарном столе»). Это название требует пояснения. Обитатели средневекового английского замка ели за одним длинным столом, примерно посередине которого стояла большая солонка. Она как бы отделяла почетные места, где сидел хозяин и его важные гости, от мест слуг, солдат и гостей второго сорта на другом конце стола; места эти называли, соответственно, «перед солью» и «за солью» («above and below the salt»). Кушанья, как легко догадаться, подавали сперва на тот конец, что «перед солью», а «за соль» доходило то, что оставалось.
Так вот, по мысли Таппан, наземная биота планеты всегда занимает место «во главе стола», а океанская – оказывается «за солью». Необходимые для жизнедеятельности фитопланктона биогены (прежде всего азот и фосфор) поступают с суши, за счет эрозии. Наземная растительность стремится подавить эрозию и оставить как можно больше биогенов для собственного потребления (вспомните о замыкании геохимических циклов). Таким образом, все заметные эволюционные успехи наземной растительности замедляют поступление биогенов в океан, что со временем вызывает вымирание соответствующих групп фитопланктона. Но фитопланктон – это основа первого трофического уровня любых морских экосистем, поэтому дальше разваливается вся цепь. Однако следует помнить, что инерция такой системы огромна и что между «прогрессивным изменением» в наземной растительности и вызванным им «фитопланктонным шоком» в океане проходит ощутимый срок.
Подойдя с этих позиций к пермо-триасовому и мел-палеогеновому морским вымираниям, мы увидим, что в поздней перми и в позднем мелу действительно произошли два серьезных изменения в структуре наземной растительности, прямо влияющие на регуляцию потока биогенов с суши. В перми высшая растительность впервые начинает заселять водоразделы, тормозя их прежде беспрепятственный размыв; этому способствует возникновение засухоустойчивых голосеменных (типа вольциевых) с пикноксилической древесиной и мелкими, сильно склеротизованными листьями (а также, как мы помним, появление насекомых с водными личинками – см. главу 9). В позднем мелу появляется трава, образующая дернину, – наиболее эффективный из известных подавителей эрозии.
Эта гипотеза вполне проверяема. Логично предположить, что когда на суше возникла наземная растительность, это тоже должно было соответствующим образом повлиять на характер стока. Суша покрылась лесами где-то к середине девонского периода (см. главу 8). А в конце девона в море действительно происходит фран-фаменское вымирание – самое крупное после пермо-триасового и мел-палеогенового!