Все перечисленные выше группы, «определяющие лицо» тропических экоситем (и опылители, и термиты, и муравьи), эволюционно очень молоды и не характерны для раннекайнофитных сообществ. Это и привело Жерихина к заключению, что тропические сообщества – наравне с бореальными – принадлежат к числу самых молодых на Земле; судя по всему, они возникли не раньше эоцена, а окончательное их формирование произошло уже в неогене.
С уверенностью говорить о времени формирования гилейных экосистем трудно из-за очень слабой палеонтологической изученности современной тропической зоны. Что же касается бореальных сообществ – например, криофильных (холодолюбивых) лесов с доминированием хвойных, типа тайги, – то они, по всей видимости, сложились в палеогене как высотный пояс в горах, а при неогеновом похолодании широко распространились на равнинах Северного полушария. В палеонтологической летописи они впервые достоверно появляются в верхнем миоцене Канадского архипелага (остров Миен); их состав напоминал современные лесотундровые редколесья.
Еще одна характерная для кайнозоя черта – травяные биомы (типа степей и саванн), существование которых основано на коэволюции злаков и крупных травоядных млекопитающих: при ряде условий (в засушливых и малоплодородных районах) животные способны воспрепятствовать восстановлению лесной растительности. В доэоценовые времена, судя по пыльцевым спектрам, сообществ такого типа на Земле не было. По мнению Жерихина (1993), эти биомы первоначально представляли собой сериальные сообщества, ведущие к соответствующим древесным климаксам (стадии с господством трав – например, луга – имеются во всех современных сукцессионных системах), однако затем возникла уникальная ситуация: фитофаги, изымая прирост биомассы (в современных степях это изъятие доходит до 60% растительной продукции – абсолютный рекорд), оказались способны остановить экогенетическую сукцессию на безлесой стадии. Дальнейшая коэволюция млекопитающих и трав привела к стабилизации этих своеобразных «зоогенных климаксов»: необходимый для климаксного сообщества нулевой баланс по органике обеспечивается в них именно деятельностью животных-фитофагов. К основным чертам, отличающим эти травяные климаксы и от сериальных травяных сообществ, и от лесов, относятся: исключительно высокая продуктивность, очень быстрый оборот органического вещества и наличие легко мобилизуемого запаса этой органики в необычайно плодородных почвах (типа черноземов).
Известно, что основой существования этих сообществ является замечательная коадаптация доминирующих в растительном покрове злаков и фитофагов: злаки не имеют эффективной защиты от поедания (вроде колючек, ядовитости и т.п.), зато способны компенсировать обгрызание надземных частей резким ускорением их регенерации. Поэтому регуляция в таком сообществе достигается предельно просто: при снижении пресса фитофагов первичная продукция тоже автоматически снижается, и наоборот. Основную роль в пастбищных цепях этих экосистем играют млекопитающие, прежде всего различные копытные (потомки кондиляртр – рис. 55, а) и хищные (потомки креодонтов – рис. 56, а); об эволюции сообщества позвоночных – чуть далее.
Иное дело детритные цепи. Возникновение стабильных травяных сообществ было бы просто невозможно без появления целого комплекса насекомых-копрофагов, перерабатывающих огромную массу экскрементов травоядных.[73] В более древних (палеогеновых) лесных сообществах, где млекопитающие не достигали такой биомассы, как пастбищные копытные, с этой работой справлялись низшие навозники. Важнейшие же современные группы копрофагов (высшие навозники и навозные мухи) в заметных количествах появляются в захоронениях с конца олигоцена. В это же время складывается и не менее важный комплекс насекомых-некрофагов (мясные мухи и жуки-мертвоеды), ответственный за эффективную переработку трупов травоядных млекопитающих.
Сообщество наземных позвоночных в кайнозое развивалось независимо на трех разобщенных территориях, фаунистические контакты между которыми практически отсутствовали. Австралия (с ее сумчатыми и однопроходными) изолирована и поныне, а Южная Америка сохраняла свою обособленность от остальной суши вплоть до плиоцена, когда возник Панамский перешеек. Отсюда и проистекает современное разделение мира на три зоогеографические области: Нотогея (Австралия), Неогея (Южная Америка) и Арктогея (Евразия, Африка и Северная Америка). Так вот, по мнению Жерихина (1993), во всех этих трех областях травяные биомы возникали независимо, на базе совершенно различных комплексов крупных млекопитающих; собственно говоря, есть серьезные основания полагать, что млекопитающие по-настоящему вышли в крупный размерный класс лишь в травяных биомах.
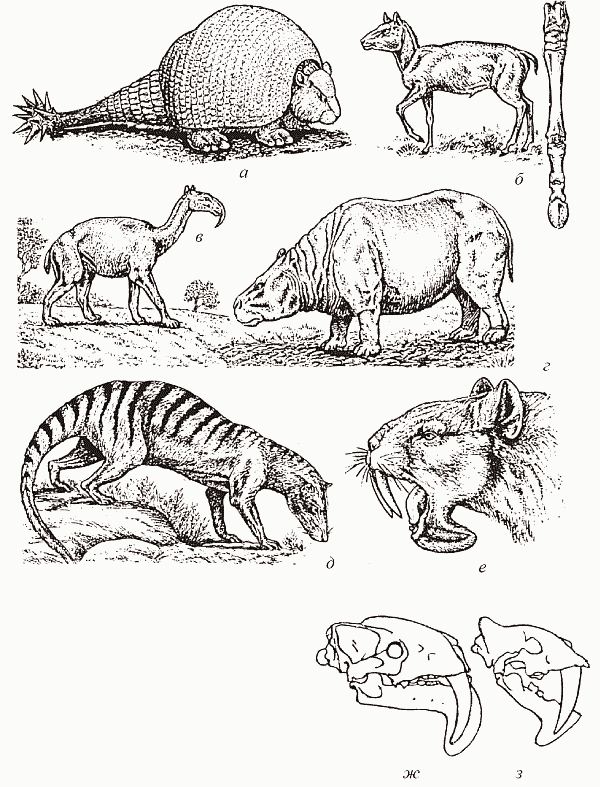
Раньше всего (в среднем эоцене) этот процесс начался в Южной Америке. Там среди исходно листоядных «южноамериканских копытных»[74] возникают первые травоядные формы, а также появляются гигантские травоядные броненосцы-глиптодонты, смахивающие на небольшой танк (рис. 54, а). В среднем же эоцене в Южной Америке впервые обнаруживаются и пыльцевые спектры с высоким содержанием пыльцы злаков, палеопочвы степного типа, а также фоссилизированные навозные шары, принадлежащие жукам-навозникам. Позднее, в олигоцене и особенно в миоцене, здесь возникает в высшей степени своеобразный комплекс пастбищных травоядных. Он включал неполнозубых (глиптодонтов и наземных ленивцев), «южноамериканских копытных» (различные литоптерны демонстрирует сильное конвергентное сходство частью с лошадьми, частью с верблюдами, у пиротериев много общего со слонами, а среди нотоунгулят были формы, схожие и с носорогами, и с бегемотами, и с кроликами – рис. 54, б–г), а также гиганских кавиморфных грызунов (некоторые из этих родственников морской свинки достигали размеров носорога) и просуществовал вплоть до установления в плиоцене сухопутной связи с Северной Америкой.
Что же касается хищников, то они в древней южноамериканской фауне всегда были в дефиците. Ни один из здешних отрядов плацентарных по не вполне понятным причинам так и не дал плотоядных форм – эту роль исполняли исключительно сумчатые. Довольно разнообразные боргиениды несколько напоминали собак (а еще больше – тилацина, тасманийского «сумчатого волка»), а тилакосмилюс вполне заслуживает названия «сумчатый саблезубый тигр» и являет собой поразительный пример конвергенции с саблезубыми кошками Северного полушария (рис. 54, д–е). Дефицит маммальных хищников (на «несбалансированность» здешних фаун обратили внимание А. С. Раутиан и Н. Н. Каландадзе, 1987) привел к тому, что эту нишу заполняли самые неожиданные персонажи. Так, с палеоцена по миоцен здесь существовали себекозухии – сухопутные крокодилы с высокой и узкой мордой (предполагают, что их образ жизни напоминал современных комодских варанов), а в эоцене появились дожившие до плейстоцена фороракосы – гигантские (ростом до 3 м) нелетающие хищные птицы, принадлежащие к журавлеобразным.
В Австралии (Нотогее) формирование травяного биома началось много позже, в неогене; здесь явно сыграл роль дрейф этого континента по направлению от полюса к экватору – в результате значительная часть его территории попала в условия засушливого климата. Основу здешнего сообщества пастбищных млекопитающих составили крупные травоядные сумчатые – кенгуру и вымершие на памяти человека дипротодонты (их иногда, из-за двух крупных резцов, не слишком удачно называют «кроликами ростом с носорога»). Как и в древней южноамериканской фауне, здесь отчетливо прослеживается дефицит хищников: известны лишь два крупноразмерных хищных млекопитающих – тилацин (тасманийский «сумчатый волк») и древесный тилаколео (которого по аналогии можно назвать «сумчатым леопардом»). Недостаток маммальных хищников возмещался (опять-таки как в Южной Америке) за счет рептилий: исполинских варанов-мегаланий длиной до 7 м и сухопутных крокодилов, сходных по образу жизни с себекозухиями; хищные нелетающие птицы здесь не появились, однако некоторые из австралийских страусов, вероятно, выполняли роль падальщиков.
73
Когда в Австралию завезли домашних копытных, оказалось, что местные навозники не справляются с утилизацией их помета и накопление неразложенных экскрементов на пастбищах останавливает возобновление трав. Пришлось завозить туда европейских навозников – тогда все пришло в норму.
74
«Южноамериканские копытные» – шесть вымерших отрядов, эндемичных для этого материка. Все они, как и наши парнокопытные и непарнокопытные, ведут происхождение от кондиляртр, однако с позднего мела развивались в изоляции и сходство с различными «северными» группами приобрели независимо, т.е. чисто конвергентно.