Третий случай формирования травяного биома – Арктогея. Здесь ситуация осложняется тем, что он формируется на единой таксономической основе (кондиляртровой), но, судя по всему, независимо в Евразии и в Северной Америке. Сообщество пастбищных млекопитающих первоначально составляют непарнокопытные (тапиры, носороги в широком смысле и халикотерии[75] ) и нежвачные парнокопытные (свинообразные и верблюды); чуть позже к ним добавляются примитивные трехпалые лошади и жвачные парнокопытные (олени) (рис. 55).
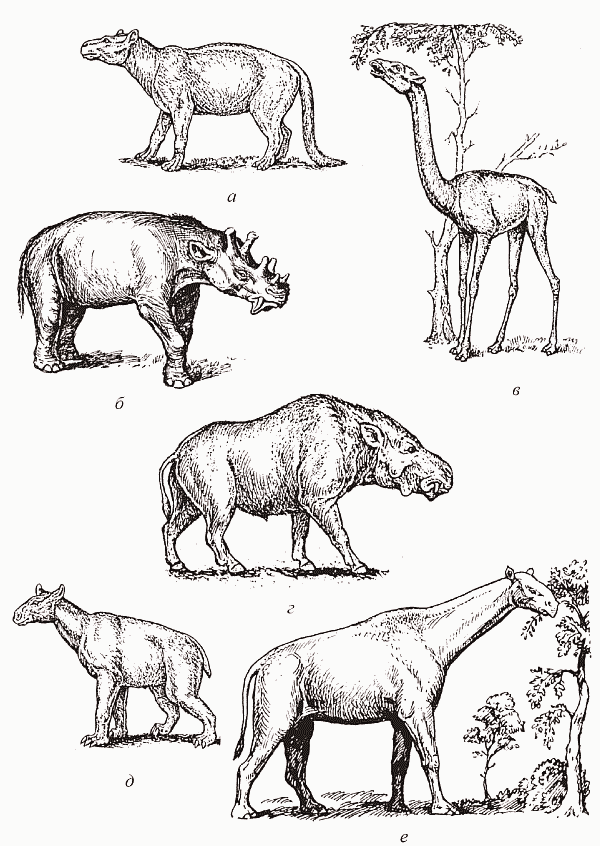
Помимо потомков кондиляртр нишу крупных растительноядных пытались освоить лишь дино-цераты – специализированые потомки каких-то крайне примитивных териевых млекопитающих (рис. 55, б), однако уже в эоцене эта группа полностью вымерла. Единство комплекса «северных» копытных достаточно высоко; самое же интересное состоит в том, что хотя практически все эти группы имеют американское происхождение (они проникали в Евразию через Берингию – область вокруг современного Берингова пролива, где тогда обсыхали обширные участки шельфа), травяные биомы с их участием в Азии начинают складываться заметно раньше, чем в Америке. В Центральной Азии саванны возникают уже в конце эоцена (появившиеся в это время гигантские безрогие носороги вроде индрикотерия – «гибрид слона и жирафа», самое крупное наземное млекопитающее, 6 м в холке – явно обитали в открытом ландшафте, а не в лесу), тогда как в Америке это происходит в олигоцене. В Африке травяных биомов, судя по всему, не существовало до миоцена; парнокопытные и непарнокопытные проникли сюда из Евразии сравнительно поздно, а эндемичные для этого континента хоботные (слоны и мастодонты) были в это время мелкими и чисто листоядными; видимо, они не могли удерживать сукцессию на безлесой стадии.
Что касается хищных млекопитающих, то на севере они, в отличие от южных континентов, были только плацентарными: сумчатые вообще существовали здесь очень недолго и так и не сумели выйти из ниши мелких насекомоядных. До того, как на севере появились специализированные плотоядные формы из креодонтов (рис. 56, а) и современных хищных (Carnivora), в этой роли подвизались своеобразные копытные – мезонихиды (рис. 56, б–в). Мезонихиды были всеядными существами (как полагают, «более плотоядными, чем кабан, но менее плотоядными, чем медведь»); они часто достигали размеров гиены, а эндрюсарх из палеоцена Монголии был крупнейшим наземным хищным млекопитающим[76] – его череп достигает в длину 85 см. Как это ни удивительно, именно от мезонихид ведут свое происхождение китообразные.
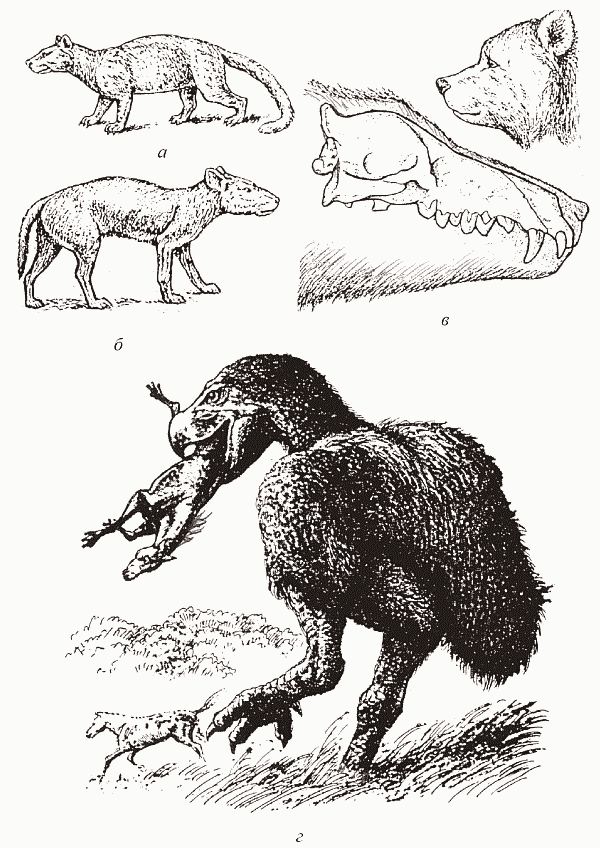
До олигоцена ситуация в травяных биомах Арктогеи и Южной Америки развивалась параллельно. И там, и там основными травоядными являлись копытные – потомки различных кондиляртр (на севере – непарно– и парнокопытные, на юге – «южноамериканские копытные»). И там, и там хищники были явно примитивнее своих жертв (на юге – сумчатые, на севере – архаичные всеядные копытные, мезонихиды) – расклад, разительно отличающий палеоген от мезозоя. И там, и там дефицит маммальных хищников возмещался за счет рептилий и птиц: сухопутных крокодилов (на юге – себекозухии, на севере – баурузухии) и гигантских нелетающих журавлеобразных (на юге – фороракосы, на севере – диатримы). Ситуация эта радикально изменилась, когда на эволюционную сцену вышли современные хищные (отряд Carnivora). Лучше всего это видно как раз по немедленному исчезновению в олигоцене всех этих «эрзац-хищников» – всеядных мезонихид, сухопутных крокодилов и диатрим, а также креодонтов (предков карнивор). Интересно, что в то же самое время исчезают и архаичные некопытные растительноядные – диноцераты.
В миоцене единство территорий Северного полушария возрастает: возникает прямой транссредиземноморский контакт между Европой и Африкой, исчезновение Тургайского моря на месте Западно-Сибирской низменности облегчает миграции между Европой и Центральной Азией, а главное – возникают открытые ландшафты в чисто лесной доселе Берингии, и эта территория превращается для степных фаун Азии и Америки из «фильтра» в «коридор». С этого времени травяной биом становится фактически единым по всей Арктогее, при этом каждая из территорий вносит свой вклад в становление его фауны, обретающей уже вполне современные черты.
Из Америки приходят травоядные (в смысле не листоядные) лошади, из Азии – полорогие (быки и антилопы)[77] , из Африки – хоботные (слоны и мастодонты); вместе с еще некоторыми группами копытных, как «новыми» (жирафы и бегемоты), так и «старыми» (носороги), они формируют так называемую гиппарионовую фауну (гиппарион – одна из трехпалых лошадей). Та же картина и с входящими в гиппарионовую фауну хищными: кошки произошли в Америке, псовые – исходно – тоже, однако стайную социальную организацию (ставшую для этой группы ключевым фактором успеха) приобрели уже в Азии, гиеновые (тогда среди них были не только падалеяды, но и активные хищники типа гепарда) – в Африке. Интересно, что кошки исходно были саблезубыми. Позже, в миоцене, возникли кошки современного типа, однако возврат к саблезубости (что, очевидно, дает преимущества при охоте на крупную добычу с особо прочной шкурой) происходил у кошачьих многократно и независимо.
В начале плиоцена (3–4 млн лет назад) самой природой был поставлен грандиозный эволюционный эксперимент: установилась сухопутная связь между Северной и Южной Америкой через Панамский перешеек, и их фауны – североамериканская (являющаяся частью Арктогеи) и южноамериканская (пребывавшая весь кайнозой в условиях островной изоляции) – вступили в прямой контакт между собой. Произошло перемешивание фаун: на севере появились сумчатые, неполнозубые (броненосцы, глиптодонты и наземные ленивцы), кавиморфные грызуны и фороракосы, а на юге – высшие грызуны (хомякообразные), непарнокопытные (лошади и тапиры), парнокопытные (свинообразные, верблюды и олени), хоботные (мастодонты) и хищные (енотовые, куньи, псовые, медведи и кошки).
Окончательные результаты Великого американского обмена (как назвал эти события Дж. Симпсон, 1983) оказались для севера и юга весьма различными. Североамериканская фауна просто-напросто обогатилась тремя экзотическими «иммигрантами» – опоссумом, девятипоясным броненосцем и древесным дикобразом, тогда как на юге произошла настоящая катастрофа, почище любых астероидных импактов: здесь полностью вымер весь пастбищный комплекс из «южноамериканских копытных», гигантских кавиморфных грызунов, хищных сумчатых и фороракосов, не выдержавших конкуренции с высшими копытными и карниворными хищниками (рис 57). Надо полагать, что судьба австралийских сумчатых и однопроходных – возникни у этого континента прямой сухопутный контакт с Азией – была бы столь же незавидной... Вообще в истории Великого американского обмена легко усмотреть прямые (и печальные) аналогии с человеческой историей: вспомним, чем обернулся «контакт» с европейской цивилизацией для древних самобытных культур доколумбовой Америки и черной Африки.
75
Это были одни из самых своеобразных копытных: пропорции конечностей у них таковы, что локомоция их была «полудвуногой», как у гориллы, а на ногах у них вторично возникли крупные невтяжные когти. Считают, что они обгладывали листву деревьев, пригибая к себе ветви при помощи «рук».
76
Именно это существо было описано И. А. Ефремовым в романе «Великая дуга» под названием «гишу – ужас ночей, пожиратель слонов».
77
Лошади (из непарнокопытных) и полорогие (из парнокопытных) являют собою две вершины в эволюции травоядности. У обоих имеется в пищеварительном тракте «бродильная камера», где симбиотические микроорганизмы дополнительно ферментируют пережеванную траву, однако у полорогих это рубец в передней части желудка, а у лошадей – слепая кишка позади него. Полорогие переваривают пищу медленно и неторопливо, извлекая из нее максимум того, что возможно, а лошади, напротив, прогоняют травяную массу через желудок с предельной быстротой. Наблюдения в африканских саваннах показали, что лошади (зебры), избирательно питаясь наиболее грубоволокнистыми травами, делают остальные типы корма более доступными для полорогих (антилоп).