Совокупность К. одного растения называется корневой системой, общую форму и характер которой определяет соотношение роста главного, боковых и придаточных К. (рис. 4 ). При преобладающем росте главного К. образуется стержневая корневая система (люпин, хлопчатник); при слабом росте или раннем отмирании главного и преобладающем развитии большого числа придаточных К. образуется мочковатая корневая система (однодольные, из двудольных — виды лютика, подорожника и др.). К. у хлебных злаков (ржи, пшеницы) проникают на глубину 1—1,5 м, у люцерны —до 10 м, у древесных растений могут углубляться на 10—12 м, хотя обычно твёрдый грунт препятствует проникновению их глубже 3—5 м; площадь, занятая корневой системой одного растения хлебных злаков, достигает в поперечнике 40—60 см, тыквы — 6—8 м, у древесных растений обычно в несколько раз превышает диаметр кроны, достигая 10—18 м. Суммарная длина К. однолетнего растения может составлять несколько километров, а общая поверхность К., учитывая поверхность волосков, во много раз превышает поверхность надземных органов. По расположению в почве в зависимости от среды обитания различают корневые системы специализированные (поверхностные или глубинные) и универсальные, равномерно развивающиеся вширь и вглубь. Например, в северной лесной зоне на подзолистых почвах, часто пересыщенных влагой, плохо аэрируемых, с трудно проницаемой подпочвой, корневая система растений на 90—95% сосредоточена в поверхностных слоях (10—15 см ). В зоне полупустынь и пустынь у одних растений имеются только поверхностные К., которые используют осадки ранней весны (эфемеры ) либо влагу дождей и конденсационную влагу, оседающую в верхних слоях почвы в ночное время (кактусы); у других К. достигают грунтовых вод (у верблюжьей колючки). Универсальные корневые системы используют в разное время влагу разных горизонтов, вследствие чего растения могут вегетировать всё лето (джузгун, саксаул, эфедра). В зонах недостаточного увлажнения ярко проявляется ярусность в расположении К. различных видов растений, произрастающих совместно на одной площади. Соотношение между К. и надземными частями растений также зависит от почвенно-климатических условий. В северной влажной зоне масса корневой системы в 5—10 раз уступает массе надземных частей. Через К. растения поглощают из почвы главным образом ионы минеральных солей, а также некоторые продукты жизнедеятельности почвенных микроорганизмов и корневые выделения др. растений. Поглощённые К. соединения азота, фосфора и серы взаимодействуют с притекающими из листьев продуктами фотосинтеза с образованием аминокислот, нуклеотидов и др. органических соединений. По сосудам ксилемы элементы в форме ионов (калий, кальций, магний, фосфор) или органических молекул (азот, сера) в результате действия корневого давления и транспирации передвигаются в листья и стебли. В К. синтезируются также алкалоиды (например, никотин), гормоны роста (кинины , гиббереллины ) и др. физиологически активные вещества. У некоторых растений, преимущественно семейство бобовых, в К. поселяются бактерии, вызывающие образование клубеньков (из паренхимы К.). К. растений выделяют в почву ионы минеральных солей, аминокислоты, сахара, мочевину, фенольные соединения и др. У одних растений (например, семейство паслёновых) К. выделяют также ауксины и вещества типа гиббереллинов, стимулирующие рост растений, у др. — студнеобразные вещества, образующие корнезащитные чехлы. Выделения К. способствуют развитию в ризосфере почвенных микроорганизмов. С возрастом растения количество выделений и численность ризосферных микроорганизмов уменьшаются. Клубеньковые бактерии играют важную роль в азотном питании растений и повышении плодородия почвы. На поверхности или внутри К. многих древесных и травянистых растений поселяются грибы, образуя т. н. микоризу ; в таких случаях корневые волоски на К. не развиваются.
На К. многих растений образуются придаточные почки, дающие надземные побеги (у корнеотпрысковых растений), У ряда растений К. служат местом отложения запасных питательных веществ (см. Корнеплоды ). У некоторых деревьев тропических лесов близ основания стволов или от ветвей отходят боковые или придаточные К. — досковидные, ходульные, столбовидные, служащие для опоры и питания. У лазящих лиан (например, у плюща) развиваются корни-прицепки. В тропиках многие эпифиты образуют придаточные воздушные К., поглощающие через многослойную поверхностную ткань (т. н. веламен) воду из атмосферных осадков. У некоторых тропических эпифитных орхидей, а также у растений семейства подостемоновых листья и стебли недоразвиты и их вегетативные органы представлены главным образом плоскими зелёными К., выполняющими функции ассимиляции. У растений, живущих на бедных кислородом почвах (например, у болотного кипариса, у мангровых деревьев и др.), имеются дыхательные К., или пневматофоры, верхушки которых расположены над почвой или водой и снабжают подземные органы воздухом. У некоторых пальм и растений семейства мареновых часть горизонтальных К. превращается в защитные колючки. К. паразитирующих на деревьях растений, например омелы, имеют вид длинных цилиндрических тяжей, расположенных в коре дерева. У растений—паразитов (заразиха, повилика и др.) и полупаразитов (марьянник, погремок и др.) корневая система развивается слабо; при этом окончания некоторых К. внедряются в виде особых присосок (гаусторий ) в тело растения-хозяина, высасывая из него питательные вещества. У некоторых растений (например, у роголистника, пузырчатки и др.) К. отсутствуют, что связано со специфическими условиями их существования. О функции К. см. также Водный режим растений , Минеральное питание растений .
К. многих растений широко используются человеком. Они имеют большое пищевое и хозяйственное значение. К., содержащие крахмал, сахара, масла, алкалоиды, гуттаперчу, красящие и др. ценные вещества, применяются в медицине и промышленности. Растения с мощно развитой корневой системой используются для закрепления подвижных песков, оврагов и эродированных почв.
Лит.: Красовская И. В., Обзор работ по морфологии и физиологии корней, «Тр. по прикладной ботанике генетике и селекции», 1928, т. 18, в. 5; её же, Закономерности строения корневой системы хлебных злаков, «Ботанический журнал», 1950, т. 35, № 4; Шалыт М. С., Подземная часть некоторых луговых, степных и пустынных растений и фитоценозов, «Труды Ботанического института АН СССР. Сер. 3, Геоботаника», 1950, в. 6; Сабинин Д. А., Физиологические основы питания растений, М., 1955; Качинский Н. А., Почва, её свойства и жизнь, М., 1956; Колесников В. А., Корневая система плодовых и ягодных растений и методы сё изучения, М., 1962; Федоров А. А., Кирпичников М. Э. и Артюшенко З. Т., Атлас по описательной морфологии высших растений, [т. 2], М.— Л., 1962; Колосов И. И., Поглотительная деятельность корневых систем растений, М., 1962; Рахтеенко И. Н., Рост и взаимодействие корневых систем древесных растений, Минск, 1963; Воронин Н. С., Эволюция первичных структур в корнях растений, «Уч. зап. Калужского педагогического института», 1964, в. 13; Бойко Л. А., физиология корневой системы растений в условиях засоления, Л., 1969; Эсау К., Анатомия растений, пер, с англ., М., 1969.
О. Н. Чистякова, Р. П. Барыкина, Д. Б. Вахмистров.
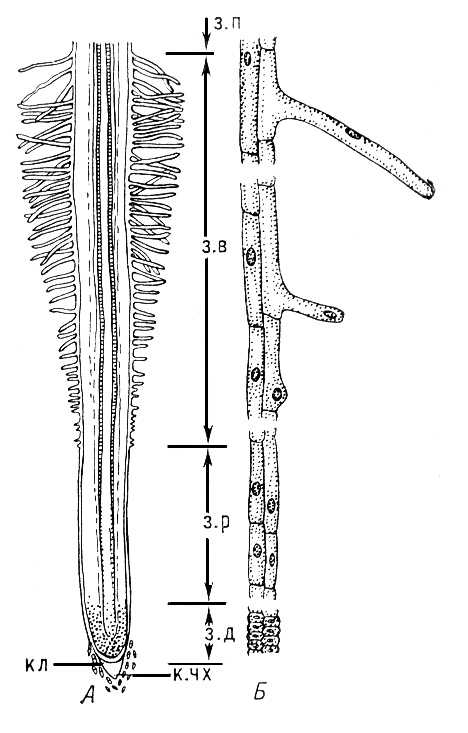
Рис. 1. Молодой корень проростка пшеницы: А — схема строения: к.чх — корневой чехлик, кл — калиптроген, з.д — зона деления, з.р — зона растяжения (роста), з.в — зона всасывания, з.п — зона проведения; Б — периферические клетки отдельных зон при большом увеличении.